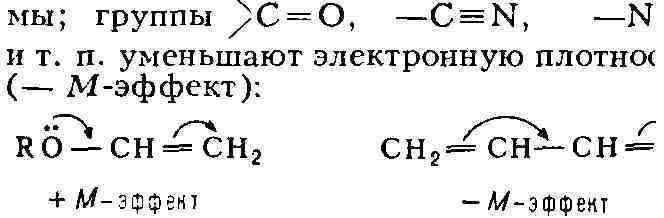![]()
![]()
|
|
|
МЁЗИЯ (Moesia), в древности страна между Ниж. Дунаем и Балканами, населённая фракийскими племенами (мёзы, геты, бессы и др.) С З. границей М. была р. Дрина, с В.- Чёрное м., на побережье к-рого в 7-6 вв. до н. э. возникли греч. колонии Одессос, Каллатия, Томы, Истрия и др. В 29-27 до н. э. была захвачена Римом. Зап. (Верхняя) М. тогда же перешла под управление римлян и не позже 15 н. э. стала рим. провинцией М.; Вост. (Нижняя) М. вошла в состав зависимой от Рима Фракии и была присоединена к провинции М. в 46. В 86 М. разделена на две провинции: Верхняя М. и Нижняя М. В кон. 3 в. Верхняя М. (М. I) вошла в диоцез М. (вместе с Македонией, Эпиром, Ахайей и о. Крит), а Нижняя М. (М. II) - в диоцез Фракия. В 4 в. М. I входила в диоцез Дакия. В 4 в. в М. оседают готы, в 6-7 вв. поселяются слав, племена. Лит.: Златковская Т. Д., Мезия в 1 и 2 вв., М., 1951. МЕЗО..., м е з... (от греч. mesos - средний, промежуточный), часть сложных слов, обозначающая ср. величину или промежуточное положение чего-либо (напр., мезодерма, мезолит). МЕЗОАТОМ, атом, в к-ром один из электронов атомной оболочки замещён отрицательно заряженным мезоном, точнее, м- мюоном, либо п- или К~-мезонами. Существование М. было предсказано амер. физиком Дж. Уилером в 1949 вскоре после открытия л~-мезонов. В 1970 было доказано существование атомов, в к-рых электрон замещён Е~-гипероном, Е~-гипероном (см. Гипероны) или антипротоном. Радиусы М. в невозбуждённом состоянии равны Гц =5,3- lO-9/mZcM, где Z - порядковый номер ядра, а т приблизительно равно отношению массы мезона к массе электрона. Наиболее простыми М. являются М. водорода. Они состоят из ядра водорода и отрицательно заряженного мезона. Их радиусы соответственно равны: rм= 2,8-10-11 см, r = 2,2-10-11см rк = = 0,8-10-и см. Такие нейтральные системы малого размера, подобно нейтронам, свободно проникают внутрь электронных оболочек атомов, приближаются к их ядрам и могут служить причиной многочисленных мезоатомных процессов: образование м е з о-молекул, катализ. ядерных реакций, перехват мезона ядрами др. атомов и т. д. В М.мезоны расположены в сотни раз ближе к ядру, чем электроны. Напр., радиус ближайшей к ядру орбиты м~ в М. свинца почти в 2 раза меньше, чем радиус ядра свинца, т. е. в М. свинца м~ осн. часть времени проводит внутри ядра. Это обстоятельство позволяет использовать свойства М. с м~ для изучения формы и размеров ядер, а также для изучения распределения электрич. заряда по объёму ядра. л- -и К- -М., кроме того, используются для изучения сильных взаимодействий элементарных частиц и распределения нейтронов в ядрах (см. Ядро атомное). Образование М. происходит, когда мезоны, получаемые в ускорителях высоких энергий, тормозятся и останавливаются в мишенях из различных веществ. Захват мезона на мезоатомную орбиту сопровождается выбросом одного из атомных электронов, обычно внешнего электрона. Напр., если пучок и~ направить в камеру с жидким водородом, то и~ теряют свою энергию в столкновениях с атомами водорода, пока их энергия не станет =<1 кэв. При этом, если они подходят близко к ядру атома водорода, они образуют с ним электрич. диполь, поле к-рого не в состоянии удержать атомный электрон, вследствие чего атом водорода теряет свой электрон, а м~ остаётся связанным с ядром (протоном, дейтроном или тритоном). Как правило, все М. образуются в высоковозбуждённых состояниях. В дальнейшем мезон переходит в менее возбуждённое состояние М., освобождая энергию в виде y-квантов(мезонное у~излучение) или Оже-электронов. На процесс образования М. влияет строение электронной оболочки молекул, в состав к-рых входит соответствующий М. Это позволяет изучать электронную структуру молекул, исследуя рентгеновское излучение М. и продукты ядерных реакций с ядром М. Это направление исследований получило название мезонной химии. Лит.: Вайсенберг А. О.,Мю-мезон, М., 1964; К im Y. N., Mesic atoms and nuclear structure, Amst. - L., 1971; Б а р х о п Э., Экзотические атомы, "Успехи физических наук", 1972, т. 106, в. 3. Л. И. Пономарёв. МЕЗОБЛACT (от мезо... и греч. blаstos - росток, зародыш, побег), средний зародышевый листок; то же, что мезодерма. МЕЗОГИППУС (Mesohippus) (от мезо... и греч. hippos - лошадь), род вымерших животных сем. лошадиных. Величиной с волка; конечности трёхпалые, все пальцы достигали земли, коренные зубы с низкой коронкой. Жили в лесах; питались в основном мягкой растительностью. Многочисленные остатки М. известны из олигоценовых отложений Сев. Америки. МЕЗОГЛЕЯ (от мезо... и греч. gloios - липкое, клейкое вещество), бесструктурное студенистое вещество у низших многоклеточных двуслойных животных (губок и кишечнополостных); выделяется эктодермой и энтодермой и заполняет пространство между ними. У медуз и гребневиков М. сильно насыщена водой (до 97,5%), В М. губок имеются амёбовидные и половые клетки, клетки, образующие скелетные элементы, и др., придающие ей характер рыхлой паренхимы. МЕЗОДЕРМА (от мезо... и греч. derma - кожа), мезобласт, средний зародышевый листок у многоклеточных животных (кроме губок и кишечнополостных) и человека. В результате гаструля-ции располагается между наружным зародышевым листком - эктодермой и внутренним - энтодермой. У первич-норотых животных (большинство беспозвоночных) М. образуется телобластическим способом - из крупных клеток - телобластов, лежащих между эктодермой и энтодермой в заднем конце зародыша и попадающих в процессе га-струляции в первичную полость тела, где они размножаются и превращаются в две мезодермальные полоски. У большинства вторичноротых животных - иглокожих, плеченогих, щетинкочелюстных, бесчерепных, круглоротых, рыб, земноводных - М. образуется энтероцель-н ы м способом: из отделяющихся участков стенки первичной кишки (ente-ron). У др. вторичноротых - пресмыкающихся, птиц и млекопитающих - благодаря вторичным изменениям процесса обособления зародышевых листков зачаток М. на стадии бластулы входит в состав первичного эктодермального слоя зародыша и лишь затем обособляется в третий зародышевый листок - М. Рис. 1. Схема развития мезодермы у кольчатых червей: 1,2, 3 - последовательные стадии; а - эктодерма, 6 - энтодерма, в - мезодермальная полоска, г - сомит, д - целом, е - спинная брыжейка, ж - мускулатура, з - кишка, и - брюшная брыжейка, к - брюшные нервные стволы, л - внутренняя сгенка целома. У плоских червей и немертин полоски М. дают начало соединит, ткани, заполняющей пространство между внутр. органами. У кольчатых червей (рис. 1) и членистоногих они расчленяются на парные сомиты со вторичной полостью тела, или целомом. За счёт стенок целома развиваются продольная мускулатура тела и выделит, органы. У разных групп позвоночных развитие М. протекает в основном сходно (рис. 2). В спинной части зародыша выделяется зачаток хорды. По обе стороны от него М. расчленяется на метамерные сомиты, к-рые сначала связаны с несегментированными брюшными отделами М.- боковыми пластинками (спланхнотомами) - узкими сегментными ножками, или нефротомами. Далее стенка каждого сомита дифференцируется на склеротом, дерматом и миотом. Склеротомы образуют осевой скелет и соединит, ткань, дерматомы -соединительнотканный слой кожи, мио-томы - скелетную мускулатуру тела. Нефротомы дифференцируются в почечные канальцы предпочки, первичной почки, а затем (у высших позвоночных) вторичной почки, а также в протоки мочеполовой системы. Спланхнотомы расчленяются на 2 листка - внутренний (висцеральный) и наружный (париетальный), между к-рыми образуется целом. Висцеральный листок примыкает к энтодерме и даёт начало гладкой мускулатуре кишечника, кровеносным сосудам и клеткам крови, а также выстилке полости тела; париетальный листок примыкает к покровам и тоже выстилает целом. В эпителии спланхнотомов возникают половые валики - зачатки половых желез. Правая и левая боковые пластинки, срастаясь над кишечником, образуют брыжейку. Рис. 2, Схема развития органов из мезодермы у высшего позвоночного (поперечный разрез зародыша): а - нервная трубка, 6 - дерматом, в - эктодерма, г - многом, д - склеротом, е - нефро-том, ж - наружный листок спланхното-ма, з - энтодерма, и - внутренний листок спланхнотома, к - эндотелий аорты, л - целом, м - хорда. Лит.: Давыдов К. Н., Курс эмбриологии беспозвоночных, П. - К., 1914; Иванов П. П., Общая и сравнительная эмбриология, М.- Л., 1937; Шмальгаузен И. И., Основы сравнительной анатомии, 4 изд., М., 1947; Ш м и д т Г. А., Эмбриология животных, ч. 1-2, М., 1951-1953; Т о к и н Б. П., Общая эмбриология, М., 1970. Т. А. Детлаф, А. В. Иванов. МЕЗОЗАВРЫ (Mesosauria), отряд вымерших пресноводных пресмыкающихся, иногда выделяемый в подкласс проганозавров. Жили в позднем карбоне -ранней перми. Дл. тела ок. 1 м. Внешний облик ящерицеобразный: голова, шея и туловище удлинённые, хвост очень длинный, сжатый с боков. Задние конечности длиннее передних; между пальцами, по-видимому, была плават. перепонка. Зубы многочисленные, игольчатые. Питались рыбой и мелкими мягкотелыми беспозвоночными. Ископаемые остатки известны из Юж. Африки и Юж. Америки. Лит.: Основы палеонтологии. Земноводные, пресмыкающиеся и птицы, М., 1964. МЕЗОЗОИ (Mesozoa), класс животных подтипа плоских червей; ранее считались промежуточной группой между простейшими и многоклеточными. 2 подкласса: дициемиды (Dicyemida) и ортонектиды (Orthonectida). Тело длиной до 5 мм, состоит из осевой клетки и имеет червеобразную форму (у дициемид) или - из скопления клеток, покрытых мерцат. эпителием (у ортонектид). М.- эндопаразиты мор. беспозвоночных. Ортонектиды живут в паренхиме турбеллярий, немертин и в полости тела, половых железах кольчатых червей, офиур, пластинчатожаберных моллюсков; дициемиды - в почках головоногих моллюсков. Жизненный цикл у М. сложный. Ортонектиды чаще раздельнополы. Развитие - со сменой бесполого и полового размножения (см. рис.). У дициемид в почках головоногих моллюсков партеногенетические поколения (нематогены) чередуются с одним гермафродитным (инфузориген). Из зигот развиваются инфузориформы (стадия распространения), к-рые выхо в воду. Ортонектид 14 видов, относящий ся к 3 родам (из 2 семейств); в СССР 7 видов (из 1 рода), обитают в Барев вом м. Дициемид-45 видов, принадле; щих к 6 родам (из 2 семейств); в СССР 12 видов (из 3 родов), обитают в да невост. морях. Схема жизненного цикла Rhopalura ophiосоrоае. А - самцы и самки, выходящие из офиуры Amphiura squa-mata; Б - оплодотворение самки самцом, В1 - самка, наполненная личинками (б1 - б3 - редукционное деление и дробление яйца внутри самки; б4 - мерцательная личинка); В - выхождение сформировавшихся личинок из самки; Г - проникновение личинок в половые щели (я) офиуры; Д - разрез через половую щель, в к-рую проникли личинки (л) паразита; Е - Е1 - личинки, проникшие в эпителий половых сумок офиуры; Ж - зачаточные плазмодии в эпителии половых щелей; 3 - то же в перитонеальном эпителии хозяина; И - молодой плазмодий; К - молодой плазмодий с моруламя (м), зародышевыми клеткамии соматическими ядрами (с), вокруг плазмодия видны ядра тканей хозяина; Л и Л1 - созревшие мужские и женские плазмодии с самцами ) и самками. Лит.: Руководство по зоологии, т. М.- Л., 1937; Б о го л е п о в а И. И., С ременная система дициемид, "Параэитол( ческий сборник", 1963, т. 21. И. И. Боголет. МЕЗОЗОЙСКАЯ ГРУППА (ЭРА) мезо... и греч. zoe - жизнь), предпоследняя группа систем стратиграфич. шкалы и соответствующая ей эра геол. истории Земли. Охватывает интервал времени примерно от 230 до 67 млн. лет назад. Длительность М. э. ок. 163 млн. лет. М. э. впервые была выделена в 1841 англ, геологом Дж. Филлипсом. Подразделяется на 3 системы (периода): триасовую систему (период), юрскую систему (период) и меловую систему (период). М. э. является временем формирования осн. контуров совр. материков и, вероятно, большинства впадин океанов (кроме Тихого, к-рый, вполне возможно, существовал раньше). Характеризуется растительностью, состоящей в основном из папоротников и голосеменных, и фауной с преобладанием рептилий среди позвоночных; в то же время является эрой возникновения покрытосеменных растений, млекопитающих и птиц. Общая характеристика. В конце палеозойской эры все древние платформы были приподняты над уровнем моря и опоясаны складчатыми горными системами, образовавшимися в результате герцинской складчатости. Восточно-Европейская и Сибирская платформы соединялись вновь возникшими горными системами Урала, Казахстана, Тянь-Шаня, Алтая и Монголии; сильно увеличилась площадь суши за счёт формирования горных областей в Зап. Европе, а также по краям древних платформ Австралии, Сев. Америки, Юж. Америки (Анды). В Юж. полушарии существовал огромный по площади древний материк Гондвана. Т. о., в конце палеозойской эры материковые блоки земной коры занимали
огромные пространства. С наступлением мезозоя началось их опускание,
сопровождавшееся трансгрессиями моря. Материк Гондвана раскололся и распался на
обособленные материки: Африку, Юж. Америку, Австралию, Антарктиду и массив
Индостанского п-ова. Начиная с юрского периода мор. воды затопили огромные
площади древних платформ (Восточно-Европейской, Индостанской,
Южно-Американской) и только что закончившие формирование складчатые области,
превратившиеся в фундамент молодых платформ (Западно-Сибирской, Скифской,
Туранской и др.). В пределах Юж. Европы и Юго-Зап. Азии начали формироваться
глубокие прогибы -геосинклинали Альпийской складчатой области. Такие же
прогибы, но на океанич. земной коре возникали по периферии Тихого ок.
Трансгрессия моря, расширение и углубление геосинклинальных прогибов продолжались
в течение мелового периода. Только в самом конце мела начинается поднятие
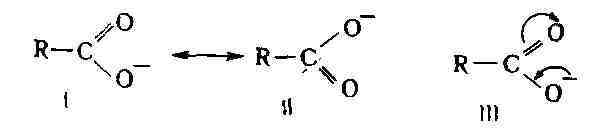
материков и сокращение площади морей. Органический мир. Преимущественно аридные условия перми и начала триаса сменились в мезозое всё более увеличивающейся влажностью климата. Обильная растительность каменноугольного периода с расцветом древовидных плауновых (лепидодендроны, сигиллярии), гигантских каламитов, кор-даитов и др. групп растений вымерла в эпоху засушливых условий перми. В М. э. происходит обновление флоры и широкое развитие растительного покрова на больших площадях материков. В триасе на материках ещё господствовали обширные аридные климатич. зоны с бедной растительностью, в к-рых в условиях равнин и озёрных водоёмов отлагались красноцветные песчано-глинистые породы с гипсом. В умеренных климатич. поясах значит, площади были покрыты лесами из хвойных (Voltzia и др.), хвощовых, папоротников, древовидных плауновых (Pleuromeia), потомков кордаитов - юкки (Vuceites). В заболоченных пространствах лесов образовались торфяные залежи, из к-рых затем возникли слои ископаемого угля и угленосные толщи (в СССР - на Урале, в Забайкалье; за рубежом - в Корее, Японии, Индии, Юж. Африке, Австралии). На Южно-Американской платформе (басе. р. Парана) и в Тунгусской впадине Сибирской платформы в триасе происходило накопление мощных туфовых и лавовых толщ, связанных с многочисл. вулканами. В юрском периоде морем были захвачены обширные пространства Европы, Зап. и Вост. Сибири, Сев. Африки и вост. побережья Тихого ок. В пределах материков образовались значит, равнинные пространства, орошаемые реками, покрытые в условиях умеренного и тропич. влажного климата пышным растит, покровом и занятые обширными озёрами и болотами. В них происходило накопление торфа, к-рый преобразовался затем в пласты ископаемого угля. В составе растит, покрова тропич. и субтропич. поясов наибольшее распространение имели голосеменные растения - хвойные, гинкговые, беннет-титовые и саговниковые (цикадовые). Среди споровых растений преобладали папоротники. Хвощовые и плауновые стали играть подчинённую роль. В сев. умеренном поясе доминировали хвойные, гинкговые и чекановскиевые леса. В триасе закончился век гигантских амфибий - стегоцефалов и уже к концу триаса преобладающую роль в фауне позвоночных получили рептилии. Гигантские рептилии достигли особенного развития в юрском и меловом периодах. Они приобрели значит, разнообразие и разделились на водных ящеров (плезиозавры и ихтиозавры), наземных ящеров - динозавров (игуанодонты, трахидонты, стегозавры и др.) и летающих ящеров (птерозавры). Особенно благоприятной средой для их развития были обширные тропич. леса и озёрно-болотные впадины, в к-рых они питались водяными растениями. В юре появились мелкие млекопитающие и первые зубастые птицы -археоптериксы. В середине мелового периода произошло сильное изменение в составе растительности. Покрытосеменные растения, первые представители к-рых появились в начале мела, к середине мела заняли господств, положение, к-рое они сохраняют до наст, времени. Влаголюбивая юрско-раннемеловая растительность постепенно заняла подчинённую роль, хотя реликты этой флоры существуют до сих пор в нек-рых тропич. и субтропич. областях (напр., в Н. Зеландии). В составе меловой флоры покрытосеменных преобладали платаны, лавры, фикусы, магнолиевые, бобовые и др. Среди хвойных были распространены сосновые, тиссо-вые, секвой, таксодиум и др. Беннеттиты к концу мела вымерли, из гинкговых остался один вид. Папоротники и саговниковые стали играть подчинённую роль. В тропич. и умеренных поясах произрастали обширные леса и продолжалось накопление торфяных залежей, давших начало угольным пластам. Развитие покрытосеменных в середине мелового периода содействовало распространению насекомых (опылителей), а это, в свою очередь, привело к широкому развитию класса птиц, а затем и млекопитающих, к-рые вытеснили пресмыкающихся. В конце М. э. крупные рептилии (динозавры) вымерли. В течение М. э. значительно изменился также состав флоры и фауны морей. Палеозойские роды и виды полностью исчезли в начале триаса и заменились новыми. Получили развитие пластинчатожаберные и брюхоногие моллюски, а пле-ченогие, преобладавшие в палеозое, отошли на второй план. Из головоногих достигли расцвета аммониты и белемниты, из иглокожих - мор. ежи и мор. лилии. Среди рыб развились и приобрели господств, положение костистые рыбы. В конце М. э. вымерли гигантские мор. рептилии (ихтиозавры), из беспозвоночных - аммониты, белемниты и др. Полезные ископаемые. К отложениям М. г., помимо залежей бурых и кам. углей (Азиат, часть СССР, Китай, США), приурочены месторождения нефти, осадочных жел. руд (СССР, Франция), бокситов (СССР, Франция, Венгрия, Румыния и др.), залежи фосфоритов и кам. соли (СССР). С интрузивными породами складчатых областей Тихоокеанского пояса связаны рудные месторождения золота (Аляска, Калифорния, Верхоянье), серебра, меди, свинца, цинка, олова. Лит.: Страхов Н. М., Основы исторической геологии, 3 изд., ч. 1 - 2, М.- Л., 1948; Жинью М., Стратиграфическая геология, пер. с франц., М., 1952; Криштофович А. Н., Палеоботаника, 4 изд., Л., 1957; Палеозойские и мезозойские флоры Евразии и фитогеография этого времени, М., 1970. М. В. Муратов, В. А. Вахрамеев. МЕЗОЗОЙСКИЕ ЭПОХИ СКЛАДЧАТОСТИ, эпохи интенсивного проявления складчатости, горообразования и гранитоидного интрузивного магматизма, происходившие в течение мезозойской эры. Наиболее интенсивно проявились по периферии Тихого ок. (в Вост. Азии, в Кордильерах и Андах), где носят назв. Тихоокеанской складчатости. Начальная тектонич. эпоха мезозойской эры - раннекиммерийская (индосинийская) - относится к концу триаса - началу юры; её проявления отмечены в Индокитае, на С.-В. Иранского нагорья, на п-овах Мангышлак и Таймыр, в сев. Добрудже и нек-рых р-нах Кордильер Сев. Америки. Следующая за ранней позднекиммерийская тектонич. эпоха, известная также под назв. андийской, невадийской, колымской, арауканской, является гл. эпохой формирования структур Верхояно-Чукотской обл., Монголо-Охотской складчатой системы, центр, части Кордильер Сев. и Юж. Америки и нек-рых др. областей. Она проявилась в конце юры - начале мела. Новое оживление тектонич. движений приходится на середину и особенно на конец мела - начало палеогена - ларамийская эпоха, когда формировалась структура Скалистых гор, зап. части Корякского нагорья, п-ова Камчатка, Сихотэ-Алиня, о. Суматра и др. Вне геосинклинальных систем мезозойский тектогенез проявился поднятиями окраинных частей платформ (особенно Сибирской и Южно-Китайской), возобновлением магматич. деятельности (кислый вулканизм, интрузии гра-нитоидов на В. Азии). Мезозойский тектогенез сопровождался образованием мно-гочисл. месторождений цветных металлов (меди, молибдена, олова, вольфрама и др.), а также золота (Тихоокеанский пояс, Монголо-Охотская система, активизированные части обрамляющих платформ и отчасти Средиземноморский пояс). Нек-рые исследователи объединяют М. э. с. и собственно альпийскую эпоху складчатости в один альпийский цикл тектогенеза. (Карту см. на стр. 7.) Лит.: Тектоника Евразии, под ред. А. Л. Яншина, М., 1966; Кордильеры Америки, пер. с англ., М., 1967; Кинг Ф., Вопросы тектоники Северной Америки, пер. с англ., М., 1969; X а и н В. Е., Региональная геотектоника, М., 1971. В. Е. Хаин. МЕЗОКАРПИЙ (от мезо... и греч. karpos - плод), межплодник, промежуточный слой околоплодника у растений. МЕЗОКЕФАЛИЯ (от мезо... и греч. kephale - голова), среднегодовое т ь, форма головы человека, характеризующаяся ср. величинами головного указателя (от 76,0 до 80,9). МЕЗОКЛИМAT (от мезо... и климат), то же, что местный климат. МЕЗОЛИТ (от мезо... и греч. li'thos -камень), эпоха каменного века, переходная между палеолитом и неолитом. Переход от палеолита к М. в основном совпал со сменой плейстоцена голоценом, характеризующимся совр. климатом, растительностью и животным миром. Дата М. Европы (установлена радиоуглеродным методом) -10-7 тыс. лет назад (в сев. р-нах он продолжался до 6-5 тыс. лет назад), М. Бл. Востока -12-9 тыс. лет назад. Для мезолитич. культур мн. территорий характерны миниатюрные кам. орудия - микролиты. Употреблялись оббитые рубящие орудия из камня - топоры, тёсла, кирки, а также орудия из кости и рога - наконечники копий, гарпуны, рыболовные крючки, острия, кирки и др. Распространились лук и стрелы, разнообразные приспособления для рыболовства и охоты на мор. зверя (долблёные челны, сети). Глиняная посуда появилась в основном уже при переходе от М. к неолиту. Собака, к-рая, ввероятно, была приручена в позднем палеолите, широко использовалась в М.; началось приручение и нек-рых др. видов животных (свинья и др.). Основой х-ва были охота, рыболовство и собирательство (в т. ч. сбор съедобных моллюсков). Отдельные мезолитические племена (напр., племена натуфийской культуры в Палестине, 10-8 тыс. до н. э.) делали попытки искусств, выращивания злаков. Т.о., возникали предпосылки для перехода (уже на ступени неолита) к производящим формам х-ва - земледелию и скотоводству. Значит, часть мезолитич. стоянок, состоявших из неск. временных жилищ, расположена на -дюнах и торфяниках. Многие стоянки представляют собой скопления раковин моллюсков (т. н. кухонные кучи), пещерные стойбища редки. Близ нек-рых мезолитич. поселений открыты родовые кладбища. Мезолитич. культуры многочисленны и разнообразны: азильская культура и тарденуазская культура в Зап. Европе; маглемозе и эртебёлле на С. Европы, себильская культура в долине Нила, капсийская культура на С. Африки, вильтон на Ю. Африки; хоабинъская культура в Юго-Вост. Азии и мн. др. Нек-рые археологи не употребляют термин "М." и относят раннемезолитич. памятники к эпипалеолиту, а поз днем езолитические - к протонеолиту или т. н. докерамич. неолиту. Лит.: У истоков древних культур (Эпоха мезолита), М.- Л., 1966 (Материалы и исследования по археологии СССР, № 126); Б а д е р Н. О., Мезолит, в кн.: Каменный век на территории СССР, М., 1970; Clark G., World prehistory, 2 ed., Camb., 1969; La prehistoire, P., 1966 (Nouvelle Clio. L'histoire et ses problemes, № 1). П. И. Борисковский. МЕЗОМЕРИЯ (отмезо ... и греч. meros-часть), сопряжение, резонансе сопряжённых системах, характер распределения электронной плотности в молекулах, к-рый можно трактовать как частичную делокализацию связей и зарядов атомов. Так, в карбоксилат-анио-не, согласно классич. структуре, один из атомов кислорода связан с атомом углерода простой связью и несёт полный отрицат. заряд, другой соединён двойной связью и нейтрален. Такая структура может быть выражена двумя равноценными формулами I и II (см. ниже). Опыт же показывает, что оба атома кислорода равноценны, т. е. каждый из них несёт один и тот же частичный отрицат. заряд, а обе связи с атомом С имеют одинаковую длину. Т. о., истинная структура является промежуточной между I и II; она может быть изображена как резонансный гибрид канонич. (крайних) структур I и II (см. Резонанса теория) или мезомерной формулой III, в к-рой изогнутые стрелки показывают направление смещения электронов, приводящего к выравниванию зарядов и связей:
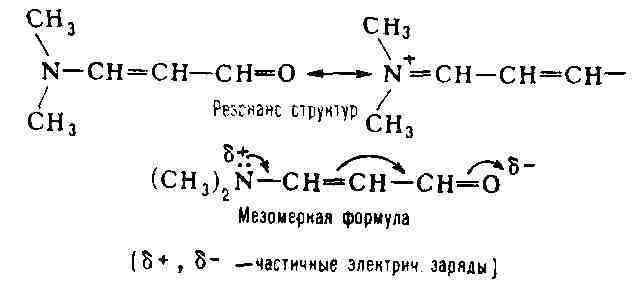
М. ярко проявляется в сопряжённых системах (см. Сопряжение связей). Обычно
она выражает состояние, промежуточное между классич. структурой и структурой
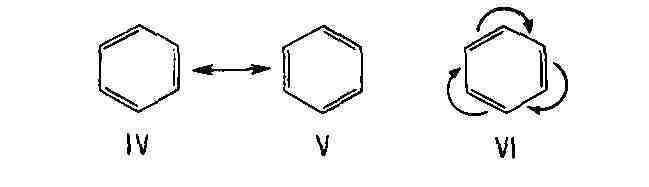
(или структурами) с полностью разделёнными зарядами, напр.: В циклич. сопряжённых системах мезомерное смещение не всегда приводит к разделению зарядов. Так, строение бензола может быть представлено как резонансный гибрид двух классич. структур Кекуле (IV и V) или же мезомерной формулой VI, отражающей равноценность всех шести атомов углерода и всех связей между ними:
Мезомерный эффект с небольшим ослабеванием передаётся по системе сопряжённых
связей (поэтому он наз. также эффектом сопряжения). Группы, несущие
неподелённую электронную пару (R2N-, RO-,
НО-, галогены), обладают положит, мезомерным эффектом (+ М-эффект) и могут
увеличивать электронную плотность остальной части систе-
Представление о М. позволяет объяснить многие свойства веществ и механизмы
реакций в органич. химии. Количеств. картина распределения электронной
плотности в молекулах может быть получена путём квантово-механич. расчётов (см.
Квантовая химия). Лит.: Несмеянов А. Н., Несмеянов Н. А., Начала органической химии, кн. 1, М., 1969. Б.Л. Дяткин. |
|
© (составление) libelli.ru 2003-2020 |