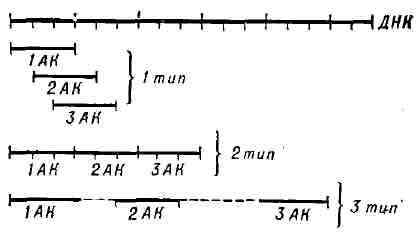
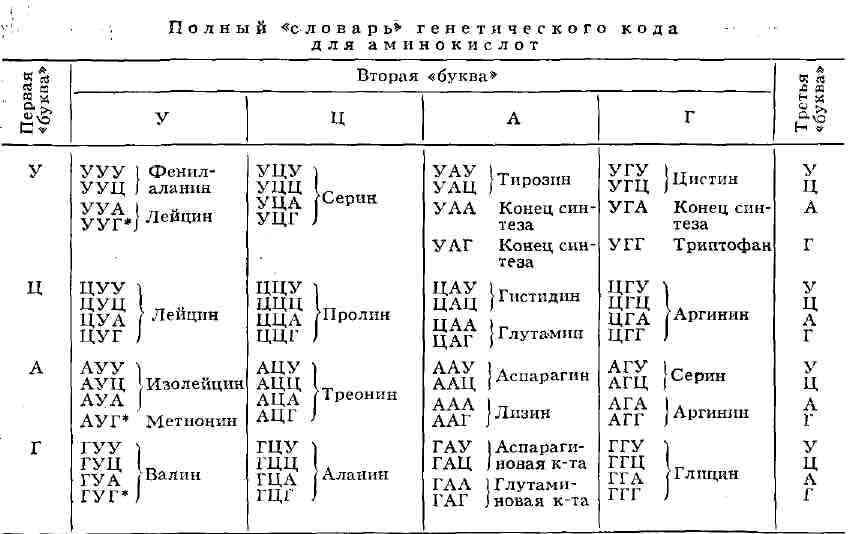
![]()
![]()
|
|
|
ГЕНЕРИРОВАНИЕ ЭЛЕКТРИЧЕСКИХ КОЛЕБАНИЙ, процесс преобразования различных видов электрич. энергии в энергию электрич. (электромагнитных) колебаний. Термин "Г. э. к." применяется обычно к колебаниям в диапазоне радиочастот, возбуждаемым в устройствах (системах) с сосредоточенными параметрами (ёмкостью С, индуктивностью L, сопротивлением R), где электрич. и магнитные поля пространственно разделены. При переходе к более высоким частотам (СВЧ и оптич. диапазон) для возбуждения колебаний необходимы системы с распределёнными параметрами. В этом случае говорят об электромагнитных колебаниях. Термин "Г. э. к.", как правило, не применяется, когда речь идёт о получении переменных токов пром. частот, получаемых с помощью электрич. машин (см. Генератор электромашинный, Переменного тока генератор). Г. э. к. осуществляется обычно либо путём преобразования энергии источников постоянного напряжения при помощи электронных приборов (вакуумных, газоразрядных и твёрдотельных), либо путём преобразования первичных электрич. колебаний в колебания требуемой частоты и формы (параметрический генератор, квантовый генератор). В зависимости от типа электронных приборов различают: ламповые генераторы (с электронными лампами), полупроводниковые генераторы (с полупроводниковыми триодами, туннельными диодами и др.), генераторы с газоразрядными приборами (тиратронами и др.). По форме колебаний, частоте, мощности и назначению различают: генераторы синусоидальных (гармонических) колебаний, генераторы колебаний специальной формы, генераторы сверхвысоких частот и т. д. Необходимые элементы генератора: источник энергии, цепи, в к-рых возбуждаются и поддерживаются колебания (пассивные цепи) и активный элемент, преобразующий энергию источника питания в энергию генерируемых колебаний. Активным элементом обычно являются электронные приборы, часто в сочетании с управляющими ими дополнит, цепями (цепями обратной связи). Если энергия, подводимая в пассивные цепи, превосходит потери энергии в этих цепях, то любой возникший в них колебат. процесс будет нарастать. Если поступление меньше потерь, колебания затухают. Энергетич. равновесие, соответствующее стационарному режиму Г. э. к., осуществимо лишь при наличии нелинейных свойств у элементов системы. При их отсутствии в системе возможен либо нарастающий, либо затухающий колебательный процесс, а генерация стационарных электрич. колебаний невозможна (см. ниже). Если цепи, в к-рых возбуждаются и поддерживаются колебат. процессы, сами по себе обладают колебат. свойствами (напр., колебательный контур или объёмный резонатор), то частота и форма генерируемых колебаний в основном определяются частотой и формой их собственных колебаний. Роль активного элемента в этом случае сводится лишь к подкачке энергии в цепи для компенсации потерь в них (включая отбор энергии потребителем). Генераторы почти гармонических колебаний. Если в генераторе с колебат. цепями потери в контуре или резонаторе малы (высокая добротность колебательной системы), то форма колебаний в них близка к синусоидальной и их наз. генераторами почти гармонических колебаний или томсоновскими генераторами. Ламповый генератор. Простейший ламповый генератор почти гармонич. колебаний состоит из колебат. контура и электронной лампы (напр., триода) с питанием и управляющей цепью (рис. 1). В контуре под влиянием случайных электрич. колебаний возникают собственные колебания тока и напряжения. Однако из-за потерь энергии в контуре колебания должны затухать. Чтобы колебания не затухали, необходимо пополнять запас колебат. энергии в контуре, напр, воздействуя на него пульсирующим током с той же частотой и с определённой фазой. Это осуществляется с помощью триода. Переменное напряжение, подводимое от контура к сетке триода, вызывает изменение его анодного тока. В результате в анодном токе появляются пульсации, к-рые при правильном подборе фазы напряжения, подаваемого на сетку лампы (цепь обратной связи), будут пополнять колебат. энергию контура. Рис. 1. Простейший ламповый генератор почти гармонических колебаний; LC - колебательный контур (С - ёмкость, L - индуктивность); Uа - анодное напряжение.
Если усилительные свойства лампы таковы, что пополнения колебательной энергии превосходят потери колебат. энергии за то же время в самом контуре, то амплитуда начальных колебаний, возникших в контуре, будет нарастать. По мере роста амплитуды колебаний усиление лампы уменьшается за счёт нелинейности вольтамперной характеристики триода и в системе установится стационарная амплитуда генерируемых колебаний. Подобные системы, генерирующие стационарные колебания, частота и форма к-рых определяются свойствами самой системы, наз. автоколебательными системами или автогенераторами, а генерируемые ими колебания - автоколебаниями. Мощность, подводимая от источника питания, расходуется не только на поддержание колебаний в контуре, но и на разогрев анода лампы электронами, бомбардирующими его при протекании анодного тока. Это обстоятельство ограничивает кпд ламповых генераторов, к-рый может всё же достигать 70-75%. Управление электронной лампой с помощью цепи обратной связи может осуществляться различными способами. Наряду с индуктивной обратной связью (рис. 1) возможна также ёмкостная обратная связь (рис. 2, а) или автотрансформаторная обратная связь (рис. 2, б). В схемах ламповых генераторов часто применяются т. н. параллельное питание анодной цепи Рис. 2. Генераторы с ёмкостной (а) и автотрансформаторной (б) обратной связью.
(рис. 2, а, б) и автоматич. смещение сетки, создаваемое сеточным током ic. Ток ic создаёт постоянное напряжение на управляющей сетке лампы, смещающее рабочую точку анодно-сеточной характеристики в область отрицат. значений, что необходимо для получения высокого кпд (рис. 3). Рис. 3. Схема лампового генератора с автоматическим смещением сетки.
Мощность ламповых генераторов - от долей вт (в измерит, и калибровочных устройствах) до десятков и сотен квт; область генерируемых частот - от десятков кгц до Ггц. Верхняя частотная граница связана, во-первых, с наличием у ламп "паразитных" ёмкостей (сетка - анод и др.), с конечным временем пролёта электронов от катода к аноду, а также с нек-рыми др. факторами (см. Электронная лампа). Нижняя частотная граница обусловлена малой добротностью колебательных контуров с низкими собственными частотами. Рис. 4. Транзисторные генераторы на плоскостных триодах с индуктивной (а), автотрансформаторной (б) и ёмкостной (в) обратной связью.
Транзисторный генератор. Другим примером генератора почти гармонич. колебаний является генератор на полупроводниковом триоде (транзисторный генератор). Здесь, так же как и в ламповом генераторе, имеется источник питания, добротный колебат. контур, а активный элемент представляет собой сочетание полупроводникового триода и цепь обратной связи. В полупроводниковых триодах (транзисторах) имеет место усиление мощности колебаний, подводимых к управляющему электроду (напр., к базе), и это позволяет, так же как и в случае электронных ламп, с помощью цепи обратной связи осуществить подкачку колебат. энергии в контур для его возбуждения и поддержания режима стационарных (незатухающих) колебаний. Существуют различные схемы транзисторных генераторов. Три варианта полупроводниковых генераторов, использующих включение транзистора по схеме с общим эмиттером, показаны на рис. 4, а, б, в. Транзисторные генераторы генерируют колебания с частотой от неск. кгц до 1010 гц с мощностями от десятых долей мвт до сотен вт. Как и в ламповом генераторе, здесь при высокой добротности контура форма колебаний близка к гармонической, а частота определяется собственной частотой колебаний контура с учётом "паразитных" ёмкостей транзистора. Отрицательное дифференциальное сопротивление. Возникновение в контуре
незатухающих колебаний можно рассматривать как результат внесения в него
нек-рого "отрицательного" сопротивления, компенсирующего
положительное активное сопротивление. В ламповом генераторе это отрицат.
сопротивление создаётся лампой в сочетании с цепью обратной связи и источником
питания. В отрицат. сопротивлении увеличение тока должно соответствовать
уменьшению падения напряжения: Эффект появления отрицательного дифференциального сопротивления возникает лишь при использовании усилительных свойств лампы или транзистора за счёт положительной обратной связи. Однако существуют приборы, в к-рых вольтамперная характеристика при
определённых условиях имеет падающий участок. Это соответствует тому, что в
нек-рой области значений U
и I имеет место
отрицат. дифференциальное сопротивление Рис. 5. Вольт-амперная характеристика с падающим участком.
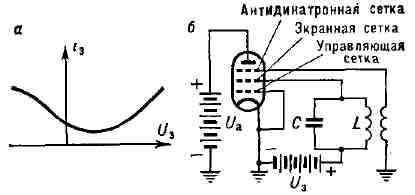
Рис. 6. a - зависимость тока экранной сетки пентода от напряжения на его антидннатронной сетке; б - схема транзитронного генератора.
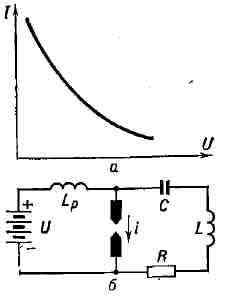
Для создания отрицательного сопротивления можно использовать электрический разряд в газах, вольтамперная характеристика которого имеет падающий участок. Напр., в определённых режимах дугового разряда с увеличением тока I возрастает темп-pa дуги, увеличивается количество ионов в разрядном промежутке и за счёт этого сопротивление промежутка падает, что приводит к уменьшению падения напряжения между электродами U. Это свойство дугового разряда использовалось в дуговых генераторах высокой частоты, применявшихся до появления ламповых генераторов (рис. 7, а, б). Рис. 7. а - вольт-амперная характеристика электрической дуги; б - дуговой генератор.
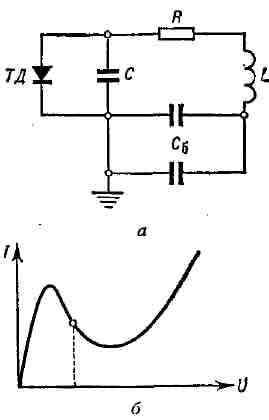
Подобным же образом может быть использована падающая характеристика туннельного диода ТД (рис. 8, а). Если рабочая точка на характеристике диода находится на падающем участке его характеристики, то это соответствует введению в колебат. контур отрицат. сопротивления. Если колебат. контур обладает высокой добротностью, то генерируемые колебания по форме близки к гармоническим и их частота определяется собственной частотой контура с учётом дополнит, ёмкости диода (подключённого параллельно основной ёмкости С, рис. 8). Рис. 8. а - генератор с туннельным диодом (ТД); 6 - вольтамперная характеристика туннельного диода.
Амплитуда установившихся колебаний будет определяться условием, чтобы
средний наклон рабочего участка характеристики (с учётом захода колебаний за
пределы наиболее крутого участка падающей характеристики) обеспечивал бы полную
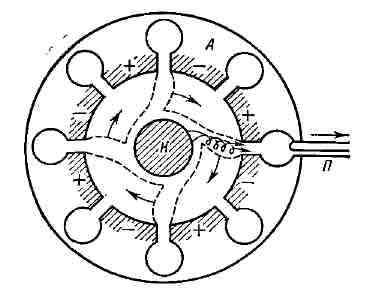
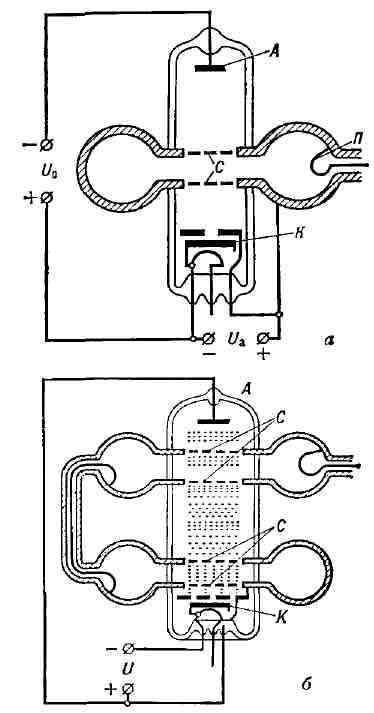
компенсацию потерь на активном сопротивлении контура, включая и полную Большинство приведённых ранее понятий (активный элемент, пассивные цепи, отрицат. сопротивление и др.) в полной мере применимо лишь к устройствам, состоящим из сосредоточенных элементов (лампа, сопротивление, конденсатор, катушка индуктивности и т. д.), размеры к-рых много меньше длины волны X. Продвижение в область СВЧ привело к созданию генераторов, представляющих собой системы с распределёнными параметрами. В этих устройствах для Г. э. к. используются различные явления, возникающие в электронных потоках в вакууме, в плазме или при прохождении тока через некоторые твёрдые тела, напр, полупроводники. В этих случаях не всегда применимо само понятие элект-рич. цепи и невозможно выделять раздельно пассивные цепи и активный элемент. Магнетронный генератор. В магнетронном генераторе колебания СВЧ возбуждаются в системе объёмных резонаторов (полости с проводящими стенками). Резонаторы расположены по окружностям массивного анода и их собственная частота v определяется диаметром полости и шириной щели, соединяющей каждую полость с общим пространством, в центре к-рого расположен катод (рис. 9). Магнитное поле, искривляя траектории электронов, движущихся от катода К к аноду А, формирует общий электронный поток, пролетающий последовательно вдоль щелей резонаторов. Магнитное поле подбирается таким, чтобы большинство электронов двигалось по траекториям, почти касающимся щелей. Т. к. в резонаторах за счёт случайных токов неизбежно возникают слабые электрич. колебания, то около щелей существуют слабые переменные электрич. поля Е. Пролетая в этих полях, электроны в зависимости от их направления относительно поля Е либо ускоряются, отбирая энергию у резонатора, либо тормозятся, отдавая часть энергии резонаторам. Электроны, ускоренные полем первого же резонатора, возвращаются на катод. Заторможенные (рабочие) электроны попадают в поле следующих резонаторов, где они также будут тормозиться, если попадают туда в "тормозящие" полупериоды электромагнитного поля. Путём соответствующего подбора скорости электронов (анодного напряжения Uа и магнитного поля Н) можно добиться того, чтобы электроны больше отдавали энергии резонаторам, чем забирали у них. Тогда колебания в резонаторах будут нарастать. Нелинейность характеристик магнетрона обеспечивает установление постоянной амплитуды генерируемых колебаний. Отбор энергии может производиться из любого резонатора с помощью петли связи П. В магнетроне источником питания является источник анодного напряжения Ua, колебат. системой - резонаторы. Роль активного элемента, обеспечивающего преобразование постоянной энергии в энергию электрич. колебаний, играет электронный поток, находящийся под действием магнитного поля.
Рис. 9. Магнетронный генератор; А - анод; К - катод; П - петля связи. Магнетроны генерируют гармонич. колебания в диапазоне частот от 300 Мгц до 300 Ггц. Кпд магнетронных генераторов достигает 85%. Обычно магнетроны используются для получения колебаний больших мощностей (неск. Мвт) в импульсном режиме и десятков квт при непрерывной генерации (подробнее см. Магнетрон).
Рис. 10. Клистронные генераторы; а - отражательный клистрон; б - двухрезонаторный пролётный клистрон; С - сетки резонатора; А - анод; К - катод. Клистронный генератор. Клистронный генератор также содержит объёмный резонатор, в к-ром колебания возбуждаются и поддерживаются электронным потоком. Поток электронов, испускаемый катодом К (рис. 10,а), ускоряется электрич. полем, создаваемым источником питания. В отражательном клистроне электроны пролетают через сетки объёмного резонатора С и, не достигая анода А, потенциал к-рого отрицателен относительно сеток резонатора, отражаются, пролетают через резонатор в обратном направлении и т. д. Если бы электроны пролетали через резонатор сплошным потоком, то в течение одного полупериода колебаний резонатора они отдавали бы резонаторам энергию, а в течение второго полупериода отнимали бы это же количество энергии у резонатора, и Г. э. к. было бы невозможно. Если же электроны влетают в резонатор отд. "сгустками", причём в такие моменты, когда резонатор их тормозит, то они отдают резонатору энергии больше, чем забирают у него. При этом электронный поток усиливает возникшие в резонаторе случайные колебания и поддерживает их с постоянной амплитудой. Т. к. группирование электронного потока в сгустки происходит за время, соответствующее неск. периодам колебаний, то протяжённость "пространства группировки" задаётся скоростью электронов и частотой генерируемых колебаний. Благодаря этому наибольшее распространение клистронные генераторы имеют в сантиметровом и миллиметровом диапазонах длин волн. Мощность клистронов невелика - от неск. мвт в миллиметровом диапазоне до песк. вт в сантиметровом. Мощность двухрезонаторных пролётных клистронных генераторов (рис. 10, б) в сантиметровом диапазоне может составлять десятки вт (подробнее см. Клистрон). Квантовые пучковые генераторы. В квантовых генераторах роль высокодобротной колебат. системы выполняют возбуждённые атомы или молекулы активного вещества. Переходя из возбуждённого состояния в невозбуждённое, они излучают порции (кванты) электромагнитной энергии, равные hv, где h - Планка постоянная, v - частота электромагнитных колебаний, характерная для данного сорта атомов. Источником энергии являются возбуждённые атомы и молекулы, а для отбора возбуждённых молекул служит сортирующая система. Напр., в молекулярном генераторе на аммиаке источником питания является источник молекулярного пучка аммиака. Объёмный резонатор, в котором находится активное вещество, осуществляет обратную связь, вызывая с помощью электромагнитного поля вынужденное излучение молекул и вложение колебат. энергии, компенсирующее потери, включая отбор энергии во вне. Аммиачный генератор работает на частоте 23,870 Ггц с весьма стабильной и узкой спектральной линией генерируемых колебаний за счёт высокой добротности квантового перехода. Высокая стабильность частоты колебаний, генерируемых квантовыми генераторами в радиодиапазоне (на аммиаке, водороде, синильной кислоте и др.), позволяет использовать их как квантовые стандарты частоты. Релаксационные генераторы. Существует широкий класс генераторов, у к-рых пассивные цепи, где возбуждаются и поддерживаются колебания, не обладают колебат. свойствами (контуры с большими потерями и др. апериодич. цепи, напр, комбинации ёмкостей С и сопротивлений R или индуктивностей L и сопротивлений R). В подобных генераторах за каждый период колебаний теряется и вновь пополняется значит, часть всей колебат. энергии. Период генерируемых колебаний при этом определяется временем релаксации (процесса установления равновесия) в этих цепях. Такие генераторы наз. релаксационными. В этом случае форма колебаний определяется совместно свойствами колебат. цепей и активного элемента и может быть весьма разнообразной - от скачкообразных, почти разрывных колебаний (напр., мультивибраторы) до колебаний, сколь угодно близких к гармоническим (RC-генераторы синусоидальных колебаний). Эта особенность релаксационных генераторов широко используется для получения электрич. колебаний спец. формы, напр, прямоугольных импульсов, пилообразного напряжения (рис. 11) и тока, а также для генерации гармонич. колебаний звуковой и сверхнизкой частот.
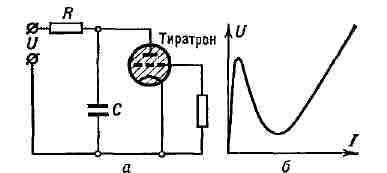
Рис. 11. Пилообразное напряжение. Тиратронный генератор пилообразного напряжения - простейший релаксационный генератор (рис. 12,а). У тиратрона напряжение зажигания выше напряжения гашения. Его напряжение U изменяется практически линейно со временем до нек-рого макс, значения, а затем достаточно быстро падает до нач. величины (рис. 11). Т. к. вольтамперная характеристика тиратрона обладает падающим участком характеристики (12, б), то процесс зарядки ёмкости С до напряжения зажигания тиратрона происходит медленно, после чего накопленный на ёмкости заряд быстро разряжается через тиратрон; напряжение на нём падает до значения, при к-ром тиратрон гаснет. При этом внутр. сопротивление тиратрона становится большим, в результате чего зарядка ёмкости С повторяется, и т. д. Период колебаний определяется временем зарядки и разрядки ёмкости, т. е. временем релаксации цепи RC.
Рис. 12. а - тиратронный генератор; б - вольтамперная характеристика тиратрона. Высокую степень линейности изменения напряжения на ёмкости можно получить, применяя вместо сопротивления R в тиратронном генераторе устройство (напр., пентод), поддерживающее постоянный ток в процессе зарядки конденсатора, или применяя отрицат. обратную связь. Частотой колебаний тиратронного генератора можно (в известных пределах) управлять, подавая синхронизирующее напряжение на сетку тиратрона. В тиратронном генераторе за период колебаний происходит полный энергообмен. Вся энергия, запасённая в конденсаторе за время зарядки, расходуется за время его разрядки через тиратрон. В этой системе нет цепей, в к-рых возможны колебат. процессы в отсутствие источников питания.
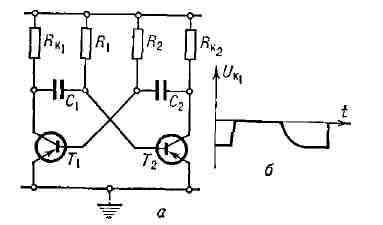
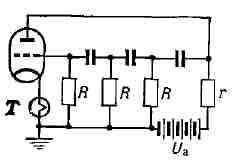
Рис. 13. Мультивибратор на транзисторах Т1 и Т2: а - схема, б - форма колебаний. Мультивибратор на электронных лампах или транзисторах представляет собой двухтактное устройство, в к-ром Г. э. к. осуществляется путём попеременной зарядки и разрядки двух ёмкостей C1 и С2 цепей RC с помощью двух взаимосвязанных транзисторов Т1 и Т2. В симметричном мультивибраторе (рис. 13,а) транзисторы Т1 и Т2 "отпираются" и "запираются" попеременно и так же попеременно происходят зарядка и разрядка ёмкостей С1 и С2. При этом резкие скачки напряжений и токов в отд. элементах схемы соответствуют быстрой смене разряда на заряд, отпиранию и запиранию транзисторов (рис. 13, б). Однако эти быстрые процессы протекают так, что запас энергии в ёмкости изменяется непрерывно. Различные варианты мультивибраторов применяются для получения периодич. напряжений различной формы, необходимых для работы электронных устройств. Период колебаний определяется временами релаксации цепей, содержащих транзисторы. Колебания возможны лишь за счёт поддержания в системе непрерывно сменяющихся процессов зарядки и разрядки в цепях RC, не обладающих собственными колебательными свойствами. RC-генератор синусоидальных колебаний также не содержит колебат. цепей. Однако за счёт выбора цепи управления активным элементом (электронной лампой, транзистором) условия Г. э. к. выполняются лишь для одного гармонич. колебания с частотой, определяемой временем релаксации цепочек RC (рис. 14). Напр., в RC-генераторе с электронной лампой термистор поддерживает усиление лампы на уровне, лишь немного превышающем критич. уровень, соответствующий условию самовозбуждения. С ростом тока растёт темп-pa термистора и увеличивается его сопротивление, что, в свою очередь, ведёт к снижению крутизны характеристики лампы за счёт возникновения отрицат. обратной связи. Т. к. работа при этом происходит практически на линейной части характеристики лампы, то условия Г. э. к. будут выполняться лишь для одной частоты. Рис. 14. RС - генератор синусоидальных колебаний; Т - термистор; r - сопротивление нагрузки.
В подобном устройстве происходит полный энергообмен за каждый период колебания. При отключении источника питания колебания исчезают, и в системе могут иметь место лишь апериодич. релаксационные процессы. С помощью RC-генератора получают гармонич. колебания в диапазоне частот от долей гц до десятков и сотен кгц. RC-генераторы широко применяются как источники эталонных колебаний. Генератор Ганна представляет собой небольшой (~100 мкм) монокристалл полупроводника, через к-рый пропускается постоянный ток. При плотностях тока, создающих в полупроводнике напряжённость поля не менее 300 кв/м (3 кв/см), в объёме полупроводника возникают нестационарные процессы, приводящие к появлению сверхвысокочастотной переменной составляющей тока, текущего через полупроводник, и к возникновению на электродах переменного напряжения СВЧ (см. Ганна эффект). В генераторе Ганна энергия источника постоянного тока преобразуется в колебат. энергию в кристалле, к-рый одновременно играет роль и колебат. системы, и активного элемента. Отсутствием высокодобротного резонатора можно объяснить немонохроматичность колебаний. Спектральная линия, соответствующая основной частоте, широка; кроме того, одновременно возбуждается большое число побочных частот. С помощью генераторов Ганна, к-рые могут применяться как маломощные гетеродины, удаётся осуществлять Г. э. к. частотой от 100 Мгц до 10 Ггц. и мощностью до 10 Мвт (при непрерывном генерировании) и сотен вт (при импульсной работе). Генераторы Ганна компактны и перспективны в микроэлектронике. Основное ограничение генерируемой мощности - нагревание кристалла при прохождении через него значит, постоянных токов. Преобразователи частоты. К ним можно отнести нек-рые типы квантовых генераторов радиодиапазона (мазеров) и оптич. диапазона (лазеров), в к-рых создание возбуждённых состояний происходит за счёт поглощения электромагнитного излучения (накачки) с частотой, существенно превышающей частоту генерируемых колебаний. Эти генераторы можно рассматривать как вторичные, преобразующие энергию колебаний накачки в колебания определённой частоты, определяемой режимом и свойствами активного вещества. Так, в радиочастотном парамагнитном мазере накачка на частоте в 10 Ггц позволяет генерировать колебания с частотой до 5 Ггц со стабильностью частоты, определяемой лишь стабильностью темп-ры и магнитного поля (см. Квантовый усилитель). В твёрдотельных лазерах на рубине или неодимовом стекле поглощение широкого спектра колебаний в области зелёной и синей части спектра приводит к генерации узкой спектральной линии с длиной волны лямбда = 6943 А (для рубинового лазера) и лямбда = 10 582 А (для лазера с неодимовым стеклом). Преобразователями частоты являются также параметрические генераторы. Параметрические генераторы радиодиапазона представляют собой резонансную колебат. систему - контур или объёмный резонатор, в к-ром один из энергоёмких (реактивных) параметров L или С зависит от приложенного напряжения или протекающего тока. При периодич. изменении одной из величин С или Z, с помощью внеш. колебаний (накачки) частоты vн в контуре могут возбуждаться и поддерживаться колебания частоты v = ½ vH. Наиболее широко распространены маломощные параметрич. генераторы с переменной ёмкостью, созданной запертым полупроводниковым диодом спец. конструкции (параметрическим диодом). Применение многоконтурных схем позволяет генерировать колебания с частотой, не связанной жёстким соотношением с частотой накачки, и тем самым осуществлять преобразование энергии исходных колебаний одной частоты в энергию колебаний требуемой частоты (см. Параметрическое возбуждение и усиление электрических колебаний). Аналогичный принцип используется для возбуждения колебаний оптического диапазона. Однако в этом случае параметрические явления носят волновой характер и осуществляются не в колебательном контуре, а в анизотропном кристалле (см. Параметрические генераторы света). Лит.: Бонч-Бруевич М. Л., Основы радиотехники, М., 1936; Xаркевич А. А., Автоколебания, М., 1954; Теодорчик К. Ф., Автоколебательные системы, М., 1952: Горелик Г. С., Колебания и волны, 2 изд., М., 1959. В. В. Мигулин. ГЕНЕТИКА. Содержание: Основные этапы и направления развития, предмет и методы генетики Основные понятия и законы генетики Генетика и эволюция Генетика и практика Основные центры генетических исследований и органы печати Литература Генетика (от греч. genesis - происхождение)- наука о законах наследственности и изменчивости организмов. Важнейшая задача Г. - разработка методов управления наследственностью и наследственной изменчивостью для получения нужных человеку форм организмов или в целях управления их индивидуальным развитием. Основные этапы и направления развития, предмет и методы генетики Основополагающие законы Г. были вскрыты чешским естествоиспытателем Г. Менделем при скрещивании различных рас гороха (1865). Однако принципиальные результаты его опытов были поняты и оценены наукой лишь в 1900, когда голл. учёный X. де Фриз, нем. - К. Корренс и австр. - Э. Чермак вторично открыли законы наследования признаков, установленные Менделем. С этого времени началось бурное развитие Г., утвердившей принцип дискретности в явлениях наследования и организации генетич. материала и сосредоточившей гл. внимание на изучении закономерностей наследования потомками признаков и свойств родительских особей. В развитии этого направления Г. решающую роль сыграл метод гибридологического анализа, сущность к-рого состоит в точной статистич. характеристике распределения отд. признаков в популяции потомков, полученных от скрещивания особей, специально подобранных в соответствии с их наследственными качествами. Уже в первое десятилетие развития Г. на основе объединения данных гибридологич. анализа и цитологии - изучения поведения хромосом в процессах клеточного деления (см. Митоз), созревания половых клеток (см. Мейоз) и оплодотворения - возникла цитогенетика, связавшая закономерности наследования признаков с поведением хромосом в процессе мейоза и обосновавшая хромосомную теорию наследственности и теорию гена как материальной единицы наследственности. Хромосомная теория объяснила явления расщепления, независимого наследования признаков в потомстве и послужила основой для понимания мн. фундаментальных биол. явлений. Под термином "ген", введённым в 1909 дат. учёным В. Иогансеном, стали понимать наследственный задаток признака. Решающий вклад в обоснование хромосомной теории наследственности был внесён работами американского генетика Т. X. Моргана (1911) и его многочисленных сотрудников и учеников, среди к-рых прежде всего следует назвать К. Бриджеса, Г. Мёллера и А. Стёртеванта. Крупной вехой в развитии Г. стало открытие мутагенного (т. е. изменяющего наследственность) действия рентгеновых лучей (сов. учёные Г. А. Надсон и Г. С. Филиппов, 1925; амер. - Г. Мёллер, 1927). Доказав резкое увеличение изменчивости генов под влиянием внешних факторов, это открытие породило радиационную генетику. Работы по радиационному и химическому мутагенезу (сов. генетики М. Н. Мейсель, 1928; В. В. Сахаров,1933; М. Е. Лобашёв, 1934; С. М. Гершензон, 1939; И. А. Рапопорт, 1943; англ. - Ш. Ауэрбах, 1944) способствовали изучению тонкой структуры гена; велико и их практич. значение для получения новых наследственно изменённых форм растений и микроорганизмов. Важное место в развитии теории гена заняли работы сов. генетиков. А. С. Серебровским была поставлена проблема сложного строения гена. В дальнейшем (1929-31) им и его сотрудниками, особенно Н. П. Дубининым, была экспериментально доказана делимость гена и разработана теория его строения из субъединиц. Г. сыграла большую роль в утверждении и развитии дарвиновской теории эволюции. Эволюционная Г. (в т. ч. популяционная Г.) исследует генетич. механизмы отбора, роль отд. генов, генетич. систем и мутационного процесса в эволюции. Фундаментальный вклад в разработку проблем Г. популяций внёс сов. генетик С. С. Четвериков (1926), объединивший в единой концепции идеи менделизма и дарвиновской теории эволюции. Развитию эволюционной и популяционной Г. особенно способствовали амер. учёный С. Райт и англ. - Дж. Холдейн и Р. Фишер, заложившие в 20-30-х гг. основы генетико-математич. методов и генетич. теории отбора. Для развития экспериментальной Г. популяций много сделали сов. учёные, гл. обр. Н. П. Дубинин и Д. Д. Ромашов, Н. В. Тимофеев-Ресовский, а также школа Ф. Г. Добржанского (США). Уже на первых этапах развития Г. внесла весьма существенный вклад в теоретич. обоснование методов селекции (работы дат. генетика В. Иогансена, 1903; швед. - Г. Нильсона-Эле, 1908). Наиболее полное выражение единство Г. и селекции нашло в трудах сов. учёного Н. И. Вавилова, открывшего гомологических рядов закон в наследственной изменчивости и обосновавшего теорию центров происхождения культурных растений. Под руководством Вавилова была проведена огромная работа по исследованию мирового разнообразия культурных растений и их диких родичей и по вовлечению их в селекционную практику. С именами Г. Д. Карпеченко и И. В. Мичурина связана разработка теории отдалённой гибридизации растений. В развитие генетических основ селекции животных крупный вклад внесли сов. генетики М. Ф. Иванов, П. Н. Кулешов, А. С. Серебровский, Б. Н. Васин и др. Сов. учёный Н. К. Кольцов (1927, 1935) впервые ясно сформулировал матричный принцип репродукции молекулярной структуры наследств, материала (хромосомы как наследственные молекулы ). Использование в качестве объектов генетических исследований микроорганизмов и вирусов (см. Генетика микроорганизмов), а также проникновение в Г. идей и методов химии, физики и математики привели в 40-х гг. к возникновению и бурному развитию молекулярной генетики. В 20-30-е гг. сов. Г. занимала ведущее место в мировой науке о наследственности и изменчивости. Начиная с 1939, а особенно после августовской сессии ВАСХНИЛ (1948) развитие сов. Г. затормозилось. С окт. 1964 вновь начался период всестороннего развития сов. Г., продолжающегося и ныне. В совр. Г. выделилось мн. новых направлений, представляющих как теоретич., так и практич. интерес. Интенсивно развивается, в частности, направление, исследующее роль генетич. аппарата в процессах онтогенеза, что привело к расширению контактов Г. с эмбриологией, физиологией, иммунологией, медициной. Важнейшей отраслью стала генетика человека и гл. обр. такой её раздел, как генетика медицинская. Разрабатываются генетич. аспекты проблемы борьбы со злокачественными новообразованиями и преждевременным старением; активно развиваются генетика поведения животных и человека и многочисленные другие отрасли Г., тесно переплетающиеся и взаимодействующие между собой. В модельных генетич. исследованиях широко пользуются специально созданными линиями животных и растений (дрозофил, мышей, крыс, кукурузы, арабидопсиса и др.), а также штаммами микроорганизмов, вирусов и культурами разных соматич. клеток. Всё шире привлекаются биохимич. и цитохимич. методы, оптическая и электронная микроскопия, спектроскопия, цитофотометрия, авторадиография, методы локального поражения клеточных органелл, рентгеноструктурного анализа. Для анализа результатов генетич. экспериментов, так же как и для их планирования, широко используются генетико-математич. методы (см. Биометрия). Основные понятия и законы генетики Совр. Г. рассматривает наследственность как коренное, неотделимое от понятия жизни свойство всех организмов повторять в ряду последовательных поколений сходные типы биосинтеза и обмена веществ в целом. Это обеспечивает структурную и функциональную преемственность живых существ - от их внутриклеточного аппарата до морфо-физиологич. организации на всех стадиях индивидуального развития. Наследственная изменчивость, т. е. постоянно возникающие изменения генотипической основы организмов, и наследственность поставляют материал, на основе которого естественный отбор создаёт многообразие форм жизни и обеспечивает поступательный ход эволюции. Одно из коренных положений совр. Г. состоит в том, что наследственная информация о развитии и свойствах организмов содержится гл. обр. в молекулярных структурах хромосом, заключённых в ядрах всех клеток организма и передаваемых от родителей потомкам. Биохимич. процессы, лежащие в основе индивидуального развития организма, осуществляются на базе поступающей из ядра информации в цитоплазматич. структурах клетки. Некоторые клеточные органеллы, в частности хлоропласты и митохондрии, обладают генетич. автономией, т. е. содержат наследственный материал. Однако в явлениях наследственности решающая роль принадлежит ядру, как это было показано, напр., в экспериментах сов. учёного Б. Л. Астаурова (см. Андрогенез). Закономерности дискретного наследования. Один из фундаментальных принципов Г. - дискретность наследственных факторов, определяющих развитие признаков и свойств. Признаки родительских особей при скрещивании не уничтожаются и не смешиваются. Развиваясь у гибридных особей первого поколения либо в форме, характерной для одного из родителей, либо в промежуточной форме, они вновь проявляются в определённых соотношениях в последующих поколениях, как это было впервые показано Г. Менделем. Скрещивая расы садового гороха, различающиеся по окраске семядолей (жёлтые и зелёные), Мендель наблюдал, что все гибридные семена первого поколения имели жёлтые семядоли; семена второго поколения, получаемые при самоопылении растений первого поколения, имели и жёлтые и зелёные семядоли; отношение между количествами таких семян равнялось 3:1. Это явление наз. расщеплением. Признак, подавляющий у гибридов первого поколения развитие контрастирующего признака (жёлтая окраска семядолей), наз. доминантным, подавляемый признак (зелёная окраска семядолей) - рецессивным. Семена второго поколения, имеющие жёлтую окраску семядолей, генетически неоднородны. Треть этих семян константна в отношении признака жёлтой окраски семядолей, растения же, развивающиеся из остальных ⅔ жёлтых семян, при самоопылении вновь расщепляются по окраске семян в отношении 3:1. Зелёные семена генетически однородны: при самоопылении растений, развивающихся из таких семян, расщепление отсутствует и все они дают только зелёные семена. Для удобства анализа явлений наследования признаков Мендель ввёл буквенную символику. Гены доминантных признаков обозначаются заглавными буквами алфавита, рецессивных - строчными. Наследственную основу организма, константного в отношении к.-л. доминантного признака, можно обозначить формулой А А; генетич. формула организма с рецессивным признаком - аа. При скрещивании организмов АА х аа возникает гибридная форма, наследственную основу к-рой можно выразить формулой Аа. Буквы Л и а обозначают соответственно гены, влияющие на развитие одного и того же признака, в данном примере - окраску семядолей. Организмы, несущие только гены, обусловливающие развитие доминантного (АА) или рецессивного (аа) признака, наз. гомозиготными; организмы, несущие и те и другие гены (Аа), - гетерозиготными. Гены, занимающие одно и то же положение в гомологичных хромосомах и влияющие на развитие одних и тех же признаков, наз. аллельными генами (см. Аллели). Явление расщепления признаков гибридных (гетерозиготных) организмов основано на том, что половые клетки (гаметы) гибридов несут только один из двух полученных ими от родителей аллельных генов (либо А, либо а). В этом состоит принцип чистоты гамет, отражающий дискретность структуры наследственного материала. Чистота гамет объясняется расхождением в мейозе гомологичных хромосом и локализованных в них аллельных генов в дочерние клетки, а числовые соотношения типов в потомстве от скрещивания гетерозиготных особей - равной вероятностью встречи гамет и заключённых в них генов при оплодотворении. Если вести анализ только по одному признаку, то обнаруживаются два типа потомков: один - с доминантным, другой - с рецессивным признаком (в отношении 3:1); если же учесть генетич. структуру организмов, то можно различить уже три типа потомков:1АА (гомозиготные по доминантному признаку), 2Аа (гетерозиготные), iaa (гомозиготные по рецессивному признаку). Проведённый Менделем анализ наследования двух разных признаков (напр., окраски семядолей и формы семян гороха) показал, что в потомстве гибридных (гетерозиготных) особей имеет место расщепление по обоим этим признакам, причём оба они комбинируются во втором поколении потомков независимо один от другого. Поскольку при расщеплении по каждому признаку возникают два типа потомков в отношении 3:1, то для случая двух независимо наследуемых признаков во втором поколении - четыре типа потомков в отношении: (3 + 1) х (3 + 1) = 9 + 3 + 3 + 1, т. е. 9/16 потомков с обоими доминантными признаками, 3/16 - с первым доминантным, вторым рецессивным, 3/16 - с первым рецессивным, вторым доминантным, 1/16 - с обоими рецессивными признаками. В случаях полного доминирования можно рассчитать соотношение типов потомков от скрещивания особей, различающихся по любому числу признаков, по формуле разложения бинома (3 + 1)n;, где п - число пар генов, по к-рым различаются скрещиваемые родительские формы. Независимость наследования, т. е. свободное комбинирование, присуща тем признакам, за развитие к-рых отвечают гены, лежащие в разных (негомологичных) хромосомах. Т. о., причина независимого наследования - в независимом расхождении негомологичных хромосом в мейозе. Последующий детальный анализ закономерностей наследования показал, что совокупность признаков организма (фенотип) далеко не всегда соответствует комплексу его наследственных задатков (генотипу), т. к. даже на одинаковой наследств, основе признаки могут развиваться по-разному под влиянием различных внешних условий. Наследственно-обусловленные признаки могут не проявиться в фенотипе либо в силу их рецессивности, либо под влиянием тех или др. факторов внешней среды. Если фенотип особи доступен непосредств. наблюдению, то о её генотипе с наибольшей полнотой можно судить на основе изучения потомков, полученных в определённых скрещиваниях. Индивидуальное развитие организмов и формирование их признаков осуществляются на основе генотипа в зависимости от условий окружающей внешней среды. Одна из основополагающих теорий Г. - хромосомная теория наследственности. Краеугольное положение этой теории состоит в том, что за развитие определённых свойств и признаков организма ответственны строго локализованные участки - гены, расположенные в хромосомах в линейном порядке. Процесс удвоения хромосом обеспечивает также удвоение генов и передачу их в каждую вновь возникшую клетку. Гены, локализованные в пределах одной хромосомы, составляют одну группу сцепления и передаются совместно; число групп сцепления равно числу пар хромосом, постоянному для каждого вида организмов (см. Кариотип). Признаки, зависящие от сцепленных (т. е. расположенных в одной хромосоме) генов, также наследуются совместно. Сцепленное наследование признаков может нарушаться в результате кроссинговера, ведущего к перераспределению во время мейоза генетич. материала между гомологичными хромосомами (см. Рекомбинация). Чем ближе друг к другу расположены гены, тем меньше вероятность их рекомбинации. На частоту рекомбинации влияют также пол особей, их физиология, состояние, а также внешние условия (темп-pa и др.). Частота рекомбинации может служить мерилом расстояния между генами. На этой основе разработаны методы определения положения генов в хромосоме и для ряда растений и животных составлены т. н. генетические карты хромосом. Для дрозофилы и кукурузы составлены также цитологич. карты хромосом, на к-рых гены локализованы в определённых, видимых под микроскопом участках хромосом. Генетич. и цитологич. карты дополняют и подтверждают друг друга. Доказано, что один ген может влиять не на один, а на мн. признаки организма (плейотропия), вместе с тем развитие каждого признака зависит не от одного, а от многих генов (полимерия). Доказано также, что функции гена и его влияние на фенотип зависят от физич. положения гена в генетич. системе (эффект положения), от совокупности остальных генов (генотипической среды) и от внешних условий. Фенотипическос выражение гена - экспрессивность, так же как и его проявление - пенетрантностъ, т. е. наличие или отсутствие контролируемого данным геном признака, могут варьировать в зависимости как от внешних условий, так и от генотипа. Под влиянием различных внешних воздействий гены могут изменяться - мутировать. К независимому мутированию способны также элементарные единицы, входящие в состав гена. Все эти факты свидетельствуют о сложности материальной структуры гена, эволюционировавшей в процессе развития жизни на Земле. После того как были вскрыты молекулярные основы организации наследственных структур и процессов, к-рые лежат в основе передачи наследств, информации в клетке (и в организме) и в поколениях клеток (и организмов), выяснилось, что гены контролируют процессы синтеза белков в клетках и что генные мутации (изменения хим. структуры генов) ведут к изменению хим. структуры белков (что в ряде случаев сводится к замене одной аминокислоты другой). Материальным носителем генетич. информации служит гигантский полимер - дезоксирибонуклеиновая кислота (ДНК), входящая в качестве важнейшего компонента в структуру хромосом всех организмов, за исключением нек-рых вирусов, содержащих рибонуклеиновую кислоту (РНК). При удвоении молекул ДНК в процессе клеточного деления дочерние молекулы при участии специфич. ферментов строятся, как на шаблоне, на материнских молекулах и точно комплементарно воспроизводят их. Записанныйв молекулярных структурах (последовательности нуклеотидов) ДНК генетический код определяет порядок расположения аминокислот в белковой молекуле. Передача информации с ДНК на синтезируемые белки осуществляется при помощи РНК. Молекулы РНК строятся на основе ДНК п комплементарны ей; вследствие этого кодирующая структура ДНК воспроизводится в молекулах РНК (см. Комплементарностъ). В клетке имеется неск. типов РНК: информационная (и-РНК), транспортная (т-РНК), рибосомная (р-РНК). Они различаются по величине молекул, структуре и функции. Порядок расположения аминокислот в белковых молекулах контролируется высокополимерной и-РНК; биосинтез белка происходит в цитоплазматич. рибонуклео-протеидных (белок + р-РНК) структурах - рибосомах - при помощи ферментов - аминоацил-р-РНК-синтетаз и энергии аденозинтрифосфата (АТФ), запасаемой в митохондриях. Транспортировка аминокислот к рибосомам осуществляется с помощью сравнительно низкополимерной т-РНК. Структура и-РНК определяет место и порядок расположения аминокислот в молекулах белка - первичную структуру белковых молекул и их осн. свойства. Ген, т. е. участок молекулы ДНК, контролирующий синтез полипептидных цепей того или иного белка, наз. структурным геном. У ряда микроорганизмов (кишечная палочка, сальмонелла), а также у фагов хорошо изучены структура и функции мн. структурных генов (цистронов); установлено, что структурные гены, контролирующие синтез ферментов определённой последовательности реакций, сцеплены в блоки (опероны). Имеются структуры (т. н. операторы), включающие синтез и-РНК структурными генами. Операторы, в свою очередь, находятся под контролем генов-регуляторов. Т. о., гены составляют сложную систему, обеспечивающую строгое согласование процессов биосинтеза в клетке и в организме в целом. В клетках в функционально активном состоянии находится лишь часть генов; активность остальных подавлена, репрессирована. В связи с закономерной сменой состояний активности генов и их репрессии меняется и спектр синтезируемых в клетке белков. Так, у человеческого плода синтезируется гемоглобин эмбрионального типа; лишь к 1 году у ребёнка гемоглобин эмбрионального типа постепенно замещается нормальным гемоглобином взрослого человека. Динамику активного и репрессированного состояний генетич. аппарата удалось наблюдать и непосредственно - с помощью микроскопич. и цитохимич. методов - на гигантских хромосомах в клетках слюнных желез личинок нек-рых двукрылых (дрозофила, хирономус). Для каждой стадии развития организма характерна строго определённая картина синтетической активности хромосом: нек-рые участки их находятся в состоянии сильной активности и синтезируют РНК, тогда как др. участки на этих стадиях развития функционально не активны, но становятся активными на др. стадиях. Оказалось, что в ряде случаев регуляторами функциональной активности генетич. аппарата являются гормоны. Проблема генетич. аспектов онтогенеза - одна из наиболее актуальных в современной биологии. Генетич. аппарат функционирует в тесном взаимодействии с внехромосомными, или внеядерными, компонентами клетки. Мн. факты свидетельствуют о важной роли цитоплазмы в осуществлении развития организма, а в ряде случаев - и в наследовании (см. Наследственность цитоплазматическая). Напр., обусловленная гибелью пыльцы мужская стерильность у кукурузы п др. растений - результат взаимодействия определённых цитоплазматич. н ядерных факторов. Давно известны факты пластидной наследственности. Свойства цитоплазмы играют большую роль при межвидовых скрещиваниях, в значит, мере определяя жизнеспособность и плодовитость гибридов. В свою очередь, свойства цитоплазмы находятся под контролем ядерного аппарата, изменение к-рого в ходе скрещиваний ведёт к изменению свойств цитоплазмы. Закономерности мутационного процесса. Наследственное разнообразие особей создаётся, с одной стороны, за счёт рекомбинации генов при скрещивании, с другой - в результате изменения самих генов, т. е. за счёт мутаций. Различают след. осн. типы мутаций: геномные, хромосомные и точковые. К геномным мутациям относится полиплоидия [увеличение числа хромосом, кратное основному, или гаплоидному (п), их числу], в результате к-рой возникают триплопды, тетраплоиды и т. д., т. е. организмы с утроенным (3п), учетверённым (4п) и т. д. числом хромосом в соматпч. клетках. Особенно большое эволюционное значение имеет амфидиплопдия, т. е. удвоение числа хромосом каждого родителя у отдалённых (межвидовых и межродовых) гибридов, что обеспечивает нормальный ход мейоза у них и восстановление плодовитости обычно стерильных гибридов. Впервые это показал Г. Д. Карпеченко (1927), получив плодовитые капустно-редечные гибриды. Мн. виды культурных растений - естественные амфидиплоиды. Так, 42-хромосомные пшеницы - сложные амфидиплоиды (гексаплоиды), несущие геномы дикой однозернянки и двух видов эгилопса - родственных пшенице диких злаков; у каждого из этих видов диплоидный набор хромосом (2 п) равен 14. Гибридное (амфидиплоидное) происхождение доказано также для овса, хлопчатника, табака, сахарного тростника, сливы и др. культурных и диких растений. Нек-рые из этих видов искусственно ресинтезированы (напр., слива - сов. генетиком В. А. Рыбиным) путём скрещивания исходных форм и последующего использования экспериментальной полиплоидии. К геномным мутациям относится также анеуплоидия (гетероплоидия), т. е. увеличение или уменьшение числа хромосом одной или неск. гомологичных пар, что ведёт к изменениям ряда признаков организма и у человека может служить причиной тяжёлых заболеваний. Мутации, относящиеся к группе хромосомных перестроек, включают различные типы реорганизаций (аберраций) хромосом и перераспределение их генетич. материала в пределах генома. Сюда относятся транслокации, т. е. взаимные обмены негомологичными участками между хромосомами; инверсии - повороты к.-л. участка хромосомы на 180o, что вызывает изменение порядка расположения генов в хромосоме; делеции - утери частей хромосом; дупликации - удвоения отд. участков хромосом. Мн. из этих изменений оказывают более или менее значит, влияние на фенотип, что свидетельствует о зависимости действия генов от их положения в геноме. Особое значение в процессе эволюции и селекции имеют точковые мутации. К группе точковых относят все мутационные изменения, при к-рых не удаётся цитологич. методами обнаружить к.-л. нарушения структуры отд. хромосом. В эту группу включают как мелкие делеции, дупликации и инверсии, так и изменения наследств, кода на молекулярном уровне (истинные генные мутации). Провести грань между этими двумя группами изменений часто не удаётся. Анализ па молекулярном уровне генных мутаций у вирусов показал, что они вызваны утерей или вставкой отд. нуклеотидов в молекуле ДНК или заменой одних азотистых оснований другими (транзиции и трансверсии) в процессе репликации (копирования) ДНК. Способность к мутациям присуща всем генам как в половых, так и в соматич. клетках организмов. Спонтанные мутации отд. генов редки, в среднем их частота равняется одной мутации на 100-200 тыс. или даже на 1 млн. генов, а иногда и ещё меньше. Это имеет определённый эволюц. смысл, т. к. создаёт стабильность наследств, системы, без чего невозможно существование самой жизни. Стабильность обеспечивается, в частности, наличием ферментов, под действием к-рых происходит репарация нарушений, возникающих в наследственных структурах. Разные гены мутируют неодинаково часто, что свидетельствует о зависимости мутабильности как от структуры гена, так и от остального генотипа. Физиологич. состояние клетки и всего организма, в частности его возраст, а также мн. условия внешней среды сильно влияют на темп мутагенеза. Большинство мутаций рецессивно; как правило, они неблагоприятно влияют на организм, делая его частично или полностью нежизнеспособным. Сильным мутагенным действием, т. е. способностью во много раз повышать частоту мутаций, обладают все виды ионизирующих излучений, ультрафиолетовые лучи и ряд хим. веществ. Все эти агенты широко используются в генетич. и селекционной практике для получения мутантных форм микроорганизмов и растений. Мутации не имеют приспособительного характера и не адекватны действующим на организм факторам: под влиянием одних и тех же воздействий могут возникать мутации разных генов; вместе с тем при различных воздействиях могут мутировать одни и те же гены. На этом основании сформулирован принцип ненаправленности мутационного процесса. Однако и при естественном, и при искусственно индуцированном мутагенезе, особенно вызванном химич. мутагенами, обнаруживается известная специфичность спектра возникающих мутаций, что связано как со своеобразием механизма действия мутагена, так и с особенностями генотипа организмов. Напр., воздействие на делящиеся клетки алкалоидом колхицином ведёт к полиплоидизации клеток, чем широко пользуются для получения новых форм растений методами экспериментальной полиплоидии. Ультрафиолетовые лучи и химические мутагены индуцируют большей частью генные мутации, тогда как нейтроны вызывают значит, процент перестроек хромосом. Обнаружены факты специфики мутирования определённых генов при различных мутагенных воздействиях. В опытах на вирусах и бактериях выявлено избирательное действие нек-рых химич. мутагенов на определённые азотистые основания, входящие в молекулу ДНК. Т. о., Г. вплотную подходит к решению проблемы управления мутационным процессом на молекулярном уровне. Однако коренная проблема совр. науки - направленное получение мутаций у сложных многоклеточных организмов - остаётся ещё не решённой. Генетика и эволюция Уже открытие Менделем закономерностей расщепления показало, что возникающие у организмов рецессивные мутации не исчезают, а сохраняются в популяциях в гетерозиготном состоянии. Это устранило одно из самых серьёзных возражений против дарвиновской теории эволюции (см. Дарвинизм), высказанное англ. инженером Ф. Дженкином, утверждавшим, что величина полезного наследств, изменения, к-рое может возникнуть у к.-л. особи, в последующих поколениях будет уменьшаться и постепенно приближаться к нулю. Г. обосновала положение, что генотип определяет норму реакции организма на среду. В пределах этой нормы условия среды могут влиять на индивидуальное развитие организмов, меняя их морфологич. и физиологич. свойства, т. е. вызывая модификации. Однако эти условия не вызывают адекватных (т. е. соответствующих среде) изменений генотипа, и поэтому модификации не наследуются, хотя сама возможность их возникновения под влиянием условий среды определена генотипом. Именно в этом смысле Г. отрицательно решила вопрос о наследовании признаков, приобретённых в течение индивидуального развития, что имело огромное значение как для утверждения дарвиновской теории эволюции, так и для селекции. Исследования показали, что природные популяции насыщены мутациями, гл. обр.
рецессивными, сохраняющимися в гетерозиготном состоянии под покровом
нормального фенотипа. В неограниченно больших популяциях при свободном
скрещивании и отсутствии давления отбора концентрация аллельных генов и
соответствующих генотипов (АА, Аа, аа) находится в определённом равновесии,
описываемом формулой англ. математика Г. Харди и нем. врача В. Вайнберга:
Генетич. исследования показали также роль мутационного процесса, изоляции, миграций, гибридизации, а также т. н. генетика-автоматических процессов в эволюционной дивергенции популяций и механизмах видообразования. Т. о., данные Г. подтвердили осн. идеи эволюционной теории Дарвина, вскрыв вместе с тем новые закономерности наследственности и изменчивости, на основе к-рых отбор создаёт бесконечно варьирующие формы живых организмов с их поразительной приспособленностью к условиям внешней среды. Генетика и практика Основой современной теории отбора и подбора служат закономерности, вскрытые общей и популяционной Г., методы оценки генетич. параметров популяций. Установив, что отбор эффективен лишь в том случае, когда он опирается на наследств, разнообразие особей в популяции, и что фенотип далеко не всегда соответствует генотипу, Г. обосновала необходимость оценки наследств. качеств и разнообразия селекционируемых организмов и вооружила селекцию соответствующими методами и практич. приёмами. Так, оценка наследств, качеств производителей по хозяйственно важным признакам их потомков, издавна практикуемая лучшими животноводами, получила на основе Г. науч. обоснование как необходимый приём селекц.-плем. работы, особенно ценный в связи с распространением метода искусств, осеменения. В основе методов индивидуального отбора у растений также лежат генетич. представления о чистых линиях, о гомо- и гетерозиготности и о нетождественности фенотипа и генотипа. Генетич. закономерности независимого наследования и свободного комбинирования признаков в потомстве послужили теоретич. основой гибридизации и скрещивания, к-рые наряду с отбором входят в число осн. методов селекции. На основе гибридизации и отбора сов. селекционерами П. П. Лукьяненко, В. С. Пустовойтом, В. Н. Мамонтовой, В. Я. Юрьевым, В. П. Кузьминым, А. Л. Мазлумовым, М. И. Хаджиновым, П. И. Лисицыным и др. созданы замечат. сорта зерновых, технич. и др. культур. Важнейшее значение для повышения эффективности селекции растений имеют закон гомологических рядов Н. И. Вавилова, его учение о генцентрах происхождения культурных растений, а также его теории отдалённых эколого-географич. скрещиваний и иммунитета. Совершенствованию методов селекции отд. видов животных и растений способствуют работы по частной Г. этих форм. Так, разведение цветных норок или каракульских овец невозможно без знания закономерностей наследования окрасок у этих животных. На основе генетич. закономерностей независимого наследования и взаимодействия генов осуществлён генетич. синтез норок с сапфировой, жемчужной и др. окрасками меха, не встречающимися в природе. Для создания новых сортов растений широкое применение получила отдалённая гибридизация, на основе к-рой получены мн. ценные сорта плодовых растений (И. В. Мичурин), пшенично-пырейные гибриды (Н. В. Цицин, Г. Д. Лапченко и др.), некоторые гибридные сорта озимых пшениц и др. Отдалённой гибридизацией успешно пользуются также в селекции картофеля, свёклы, ряда древесных культур, табака и др. Явление цитоплазматич. мужской стерильности используют в селекции кукурузы, пшеницы, сорго и др. культур. Всё большее практическое значение приобретают методы экспериментальной полиплоидии для создания хозяйственно ценных форм с.-х. растений. Этими методами созданы высокопродуктивные триплоидные гибриды сахарной свёклы, гречихи, триплоидный бессеменной арбуз, полиплоидная рожь, клевер, мята и др. Всё шире практикуется, особенно применительно к микроорганизмам, вызывание мутаций ионизирующей радиацией и химич. мутагенами. Уже созданы мутантные штаммы продуцентов ряда антибиотиков, аминокислот, ферментов и др. биологически активных веществ, во много раз превосходящие по продуктивности исходные штаммы (см. Генетика микроорганизмов). Искусств. мутагенез, применённый в селекции растений в СССР ещё в кон. 20-х гг. (Л. Н. Делоне, А. А. Сапегин и др.), ныне широко используется в селекц. работе в разных странах. На основе искусственно полученных мутантных форм созданы высокоурожайные сорта ячменя, пшеницы, риса, овса, гороха, сои, фасоли, лупина и др., уже внедрённые в произ-во. Значительно повышая наследств, изменчивость растений, методы экспериментальной полиплоидии и искусств, мутагенеза ускоряют селекционную работу и делают её более эффективной. Это, однако, не умаляет роли отбора и гибридизации. Значение старых методов выведения сортов и пород в сочетании с новыми приёмами, основанными на успехах Г., всё больше возрастает, особенно в селекции животных, где экспериментальная полиплоидия и мутагенез пока ещё не применимы. Разработка теории и методов оценки, отбора и подбора животных и растений, так же как и системы их наилучшего выращивания, остаётся важной задачей. На достижениях Г. основаны методы генетически регулируемого гетерозиса, обеспечившие получение гибридной кукурузы, урожайность к-рой на 30-40% выше исходных сортов, сорго и др. культур, а из с.-х. животных - свиней и особенно кур (лучшие гибридные куры превосходят чистопородных кур или межпородных гибридов по яйценоскости, крупности яиц, оплате корма) (см. Генетика животных и Генетика растений). Всё большую роль играет Г. в изучении наследственности человека, в предупреждении и лечении наследств, болезней (см. Генетика человека, Генетика медицинская ). Г. внесла большой вклад в познание диалектико-материалистич. картины мира, показав, что коренное свойство жизни - наследственность - основывается на сложной физико-химич. структуре хромосомного аппарата, сформировавшегося в ходе эволюции для хранения и передачи генетич. информации. Тем самым Г. дала ещё одно доказательство взаимосвязи физико-химич. и биологич. форм организации материи и единства материального мира. Г. показала, что все генетич. явления и процессы, в т. ч. явления наследств, изменчивости, детерминированы. Диалектически противоречивое единство явлений наследственности и наследств, изменчивости получило объяснение в поведении и особенностях изменения структуры хромосом и заключённых в них генов при скрещиваниях, а также в реакции генетич. материала на внешние воздействия или на условия внутриклеточной среды. Г. показала также, что гл. обр. внутреннее противоречие между наследственностью и наследств, изменчивостью, разрешаемое в процессе мутирования, рекомбинации при гибридизации и отбора, служит движущей силой эволюции. Г. подтвердила эволюц. теорию Дарвина и способствовала её развитию. Вскрыв материальность явлений наследственности, Г., в силу самой логики развития естествознания, показала, что все генетические явления и процессы подчинены законам диалектич. движения. Развивая теорию наследственности и изменчивости, сов. генетики твёрдо стоят на позициях диалектического материализма, марксистско-ленинской философии. Основные центры генетических исследований и органы печати В СССР гл. центрами исследований по Г. являются Ин-т общей генетики АН СССР, Ин-т биологии развития АН СССР, Ин-т молекулярной биологии АН СССР, Отдел химич. генетики Ин-та химич. физики АН СССР, Радиобиологич. отдел Ин-та атомной энергии АН СССР, Ин-т мед. генетики АМН СССР (все в Москве), Ин-т цитологии и генетики Сиб. отделения АН СССР (Новосибирск), Ин-т генетики и цитологии АН БССР (Минск), Ин-т цитологии АН СССР (Ленинград), Ин-т генетики и селекции пром. микроорганизмов Главмикробиопрома (Москва), Сектор молекулярной биологии и генетики АН УССР (Киев), а также кафедры генетики МГУ, ЛГУ и др. ун-тов. Организовано (1965) Генетиков и селекционеров общество Всесоюзное им. Н. И. Вавилова с отделениями на местах. Г. преподают во всех ун-тах, мед. и с.-х. вузах СССР. Генетич. исследования интенсивно ведутся и в др. социалистич. странах, а также в Великобритании, Индии, Италии, США, Франции, ФРГ, Швеции, Японии и др. Каждые 5 лет собираются междунар. генетич. конгрессы (см. Биологические конгрессы международные). Осн. печатный орган, систематически публикующий статьи по Г., - журнал Генетика АН СССР (с 1965). АН УССР издаёт журнал "Цитология и генетика" (с 1967). Статьи по Г. печатают также мн. биологические журналы, напр. "Цитология" (с 1959), "Радиобиология" (с 1961), и "Молекулярная биология" (с 1967). За рубежом статьи по Г. печатают десятки журналов и ежегодников, напр. Annual Review of Genetics (Palo Alto, с 1967), Theoretical and Applied Genetics (В., с 1929), Biochemical Genetics (N. Y., с 1967), Molecular and General Genetics (В., с 1908), Heredity (Edinburgh, с 1947), Genetical Research (Camb., c l960), Hereditas (Lund,cl920), Mutation Research (Amst., с 1964), Genetics (Brooklyn - N. Y., с 1916), Journal of Heredity (Wash., с 1910), Canadian Journal of Genetics and Cytology (Ottawa, с 1959), Japanese Journal of Genetics (Tokyo, с 1921), Genetica Polonica (Poznan, с 1960), Indian Journal of Genetics and Plant Breeding (New Delhi, с 1941). Д. К. Беляев. Лит.: История генетики - Мендель Г., Опыты над растительными гибридами, М., 1965; Морган Т., Избранные работы по генетике, пер. с англ., М.- Л., 1937; Вавилов Н. И., Избр. соч. Генетика и селекция, М., 1966; Гайсинович А. Е., Зарождение генетики, М., 1967; Рейвин А., Эволюция генетики, пер. с англ., М., 1967; Классики советской генетики. 1920-1940, Л., 1968. Учебники и руководства - Руководство по разведению животных, пер. с нем., т. 2, М., 1963; Бреславец Л. П., Полиплоидия в природе и опыте, М., 1963; Молекулярная генетика, пер. с англ., ч. 1, М., 1964; Сэджер Р. и Райн Ф., Цитологические и химические основы наследственности, пер. с англ., М., 1964; Волькенштейн М. В., Молекулы и жизнь. Введение в молекулярную биофизику, М., 1965; Актуальные вопросы современной генетики. Сб. ст., М., 1966; Бреслер С. Е., Введение в молекулярную биологию, 2 изд., М.- Л., 1966; Дубинин Н. П., Глембоцкий Я. Л., Генетика популяций и селекция, М., 1967; Алиханян С. И., Современная генетика, М., 1967; Мюнтцинг А., Генетика. Общая и прикладная, пер. с англ., 2 изд., М., 1967; Лобашев М. Е., Генетика, 2 изд., Л., 1967; Уотсон Дж., Молекулярная биология гена, пер. с англ., М., 1967; Боннер Дж., Молекулярная биология развития, пер. с англ., М., 1967; Робертис Э. де, Новинский В., Саэс Ф., Биология клетки, пер. с англ., М., 1967; Медведев Н. Н., Практическая генетика, 2 изд., М., 1968; Гершкович И., Генетика, пер. с англ., М., 1968; Хатт Ф., Генетика животных, пер. с англ., М., 1969; Дубинин Н. П., Общая генетика, М., 1970. Словари - Ригер Р., Михаэлис А., Генетический и цитогенетический словарь, пер. с нем., М., 1967. "ГЕНЕТИКА", научный журнал АН СССР. Издаётся в Москве с 1965. В журнале помещаются статьи, обзоры, сообщения по вопросам молекулярной, химич. и радиационной генетики, мутационной теории, генетики популяций, цитогенетики, взаимоотношений генотипа и среды, теории селекции, частной генетики и селекции животных, растений и микроорганизмов, экспериментальной полиплоидии, гетерозиса, отбора и гибридизации, генетики человека. К статьям даётся резюме на англ, языке. Тираж (1971) ок. 3,2 тыс. экз. Журнал переводится и издаётся на англ. языке в США. ГЕНЕТИКА ЖИВОТНЫХ, раздел генетики, изучающий наследственность и изменчивость преим. с.-х., а также домашних и диких животных. Основывается на общегенетич. принципах и положениях и использует в основном такие методы общей генетики, как гибридологический, цитологический, популяционный, онтогенетический, математико-статистический, близнецовый и др. Чаще всего у животных наблюдается независимое наследование признаков, обусловленное большим числом хромосом. Напр., диплоидное число хромосом у уток 80, у собак и кур по 78, лошадей 66, кр. рог. скота и коз по 60, овец 54, кроликов 44, свиней 40, лисиц 38, норок 30. Осн. методом изучения наследования признаков служит гибридологический анализ. Этот метод позволил выяснить характер наследования мн. морфологич., физиол. и биохим. особенностей, часто зависящих только от одной или неск. пар генов. Большое внимание уделяется генетике биохим. свойств молока, крови животных, в частности иммуногенетике, результаты к-рой используются для контроля за родословными племенных животных, уточнения их происхождения в спорных случаях и т. д. Установлена возможность с помощью изучения генов, обусловливающих биохим. свойства, вести анализ структуры пород, их линий и отродий, судить о степени однотипности пород и т. п. Продолжаются исследования коррелятивных связей этих генов с продуктивностью, плодовитостью и жизнеспособностью животных. Генетич. объяснение получили встречающиеся у животных морфологич. недостатки и недоразвитие отдельных органов. Известно, что мн. из пороков развития (бульдоговидность, карликовость и водянка головы у телят, безногость у поросят, безволосость у телят и крольчат и др.) определяются т. н. летальными и полулетальными генами. Особи - носители таких генов, или гибнут, или обладают низкой жизнеспособностью. Появление животных с такими недостатками объясняется тем, что в стадах встречаются особи, внешне нормальные и вполне жизнеспособные, но гетерозиготные по генам, определяющим эти недостатки. При скрещивании таких гетерозиготных особей друг с другом в потомстве появляются нежизнеспособные формы, гомозиготные по летальным или полулетальным генам. Летальное или полулетальное действие могут оказывать и гены, обусловливающие полезные в хоз. отношении признаки. Классич. пример этого - доминантный ген, определяющий у каракульских ягнят серую окраску - ши-рази, к-рый одновременно оказывается рецессивным в отношении жизнеспособности особей. Новым и перспективным направлением Г. ж. является генетика устойчивости к нек-рым инфекционным, инвазионным и грибковым заболеваниям. Известны генетически обусловленные различия устойчивости животных к маститу, туберкулёзу, ящуру, пироплазмозу и др. Мало изучены у животных наследственные болезни обмена веществ, хотя по аналогии с генетикой человека можно предполагать, что они также многочисленны. Развитие у животных количественных признаков - скороспелости, величины удоя, содержания жира в молоке, настрига шерсти, яйценоскости и др. - зависит от деятельности мн. систем организма. Этим объясняется сложная генетич. природа этих признаков. Установлено, что количественные признаки определяются совокупным действием мн. генов с однозначным действием. Последние могут различаться по степени доминирования, вплоть до сверхдоминантных генов, вызывающих гетерозис в первом поколении помесей. Для изучения количественных признаков пользуются математико-статистич. методами. Породы и внутрипородные группы с.-х. животных (линии, семейства и т. д.) - всегда популяции, в к-рых происходит расщепление по мн. генам. Популяц. метод позволяет изучить распространение отдельных генов в популяциях животных. В простейших случаях, при расщеплении в популяции по одному или немногим генам, параметрами, характеризующими популяции, служат частоты отдельных генов. При анализе признаков, зависящих от мн. генов, частоты отдельных генов не могут быть установлены, и тогда пользуются коэффициентом наследуемости - отношением генотипич. изменчивости количественного признака к его общей фенотипич. изменчивости. Значения коэфф. наследуемости (от 0 до 1) зависят от специфики признаков, для к-рых они устанавливаются, а также от степени вырав-ненности условий содержания и кормления и от методов разведения животных. Значение коэфф. наследуемости позволяет найти наиболее подходящие методы селекции и прогнозировать их результаты. Лит.: Брюбейкер Дж. Л., Сельскохозяйственная генетика, пер. с англ., М., 1966; Дубинин Н. П., Глембоцкий Я. Л., Генетика популяций и селекция, М.. 1967; Генетические основы селекции животных. Сб. ст., М., 1969; Серебровский А. С., Генетический анализ, М., 1970; Хатт Ф. Б., Генетика животных, пер. с англ., М., 1969. См. также лит. при ст. Генетика. П. Ф. Рокицкий. ГЕНЕТИКА МЕДИЦИНСКАЯ, раздел генетики человека, изучающий наследственные заболевания и методы их предупреждения, диагностики и лечения. Существование заболеваний, передающихся по наследству (гемофилия и др.), а также тот факт, что браки между родственниками увеличивают частоту появления в потомстве наследственных заболеваний, были известны давно. В нач. 20 в. исследовалась гл. обр. соотносительная роль наследственности и среды в происхождении не только нормальных, но и патологич. признаков человека. В России Г. м. зародилась лишь при Советской власти и значит, развития достигла в 30-е гг. В Медико-генетическом ин-те, возглавлявшемся С. Г. Левитом, успешно изучалось наследование сахарного диабета, язвенной и гипертонич. болезней и др. С. Н. Давиденков и его школа всесторонне исследовали наследств, болезни нервной системы. При изучении наследств, болезней Г. м. пользуется всеми методами генетики человека: генеалогическим (заключается в составлении родословной, что позволяет выявить соотношение между здоровыми и больными членами семьи пробанда, т. е. больного, для к-рого составляется родословная), близнецовым (см. Близнецы), цитологическим, биохимическим, иммунологическим. В 60-х гг. 20 в. большое значение приобрели цитогенетич. методы исследования (хромосом и полового хроматина) и биохим. тесты. Выяснилось, что наследств, болезни зависят от изменений в хромосомах половых клеток,- это могут быть изменения структуры генов, хромосомные перестройки (делеции, транслокации, дупликации и др.) или количественные изменения в хромосомном наборе (добавление или потеря одной или неск. хромосом). Так, нарушением числа или структуры хромосом - половых или аутосом - обусловлены синдром Тернера - Шерешевского, Дауна болезнь, синдромы Клайнфельтера и кошачьего крика, трисомии 18 (Эдвардса) и D (Патау) и др. При др. наследств, заболеваниях обнаружить видимые дефекты в хромосомном наборе не удаётся. В таких случаях, по-видимому, происходят генные мутации или неблагоприятное сочетание различных генов. Проявление нек-рых наследств, болезней зависит от средовых факторов, к-рые могут способствовать развитию наследств, предрасположения или полностью его подавить. В задачу Г. м. входит выявление соответств. условий среды (в т. ч. леч. средств, диеты и др.). Успехи в развитии Г. м. сделали возможными предупреждение и лечение ряда наследств, болезней. Один из эффективных методов такого предупреждения - медико-генетическое консультирование с предсказанием риска появления больного в потомстве лиц, страдающих данным заболеванием или имеющих больного родственника. Достижения биохим. генетики раскрыли первичные (молекулярные) дефекты при мн. наследственно обусловленных аномалиях обмена веществ, что способствовало развитию методов экспресс-диагностики, позволяющих быстро и рано выявлять больных и лечить мн. прежде неизлечимые наследств, болезни. Так, напр., подбором спец. диеты возможно предупредить развитие фенил-кетонурии (фенилпировиноградной олигофрении) и нек-рых др. наследств, болезней. В иных случаях лечение состоит во введении в организм извне веществ, не образующихся в нём в результате генетич. дефекта. Мн. генетич. дефекты исправляются своеврем. хирургич. вмешательством или педагогич. коррекцией. Во мн. странах организованы клиники и научные ин-ты, занимающиеся изучением наследственной патологии человека; в СССР - Ин-т мед. генетики АМН СССР. Ежегодно публикуется ок. 3 тыс. работ по Г. м. Вопросы Г. м. освещают журналы Acta geneticae medicae et gemello-logiae (Roma, с 1952); American Journal of Human Genetics (Baltimore, с 1949); Annals of Human Genetics (L., с 1954); Journal de genetique humaine (Geneve, с 1952); Journal of Medical Genetics (L., с 1964); Excerpta medica, Section. 22, Human genetics (Amst., с 1962). В СССР статьи по Г. м. публикуются во мн. мед. и биол. журналах (напр., в журн. Генетика, Цитология, Цитология и генетика). Е. Ф. Давиденкова. ГЕНЕТИКА МИКРООРГАНИЗМОВ, раздел общей генетики, в к-ром объектом исследования служат бактерии, микроскопии, грибы, актинофаги, вирусы животных и растений, бактериофаги и др. микроорганизмы. До 40-х гг. 20 в. считалось, что, поскольку у микроорганизмов нет ядерного аппарата и мейоза, на них не распространяются Менделя законы и хромосомная теория наследственности. С начала 40-х гг. микроорганизмы становятся объектом интенсивных геиетич. исследований. Именно на них были решены мн. кардинальные вопросы совр. генетики. Так, первое указание на то, что материальным носителем наследственности служит дезоксирибонуклеиновая кислота (ДНК), было получено в опытах на пневмококках (амер. генетики О. Т. Эйвери, К. Мак-Леод и М. Маккарти). Примерно в то же время были начаты интенсивные генетич. исследования на хлебной плесени - нейроспоре. Изучение многочисл. биохим. мутантов нейроспоры (Дж. У. Бидл и Э. Л. Тейтем, США) привело к установлению очень важного положения: один ген - один фермент (ныне это положение более точно формулируется так: один ген - одна полипептидная цепь). Генетич. исследования микроорганизмов особенно интенсивно стали развиваться после того, как амер. генетики С. Лурия и М. Дельбрюк показали на кишечной палочке (Escherichia coli), что и бактерии подчиняются мутационным закономерностям (см. Изменчивость, Мутации). Ранее существовавшее представление об адекватной, адаптивной изменчивости у бактерий возникло вследствие методич. ошибки, заключавшейся в изучении культуры как единицы изменчивости. Был предложен новый принцип изучения изменчивости у бактерий - клональный анализ, т. е. изучение потомства одной клетки - родоначальницы клона. Важной вехой в развитии Г. м. явился разработанный амер. генетиками Дж. и Э. Ледербергами метод реплик, или отпечатков, позволивший доказать, что мутации возникают у бактерий независимо от условий культивирования, и, кроме того, значительно упростивший приёмы отбора вариантов микроорганизмов с желаемыми свойствами. Оказалось, что в больших популяциях бактериальных клеток мутации возникают спонтанно. В 1946 был открыт половой процесс у бактерий (конъюгация), что позволило применить для их исследования генетический анализ. В результате установлены наличие у бактерий рекомбинации, существование у них генетич. групп сцепления и построены гепетич. карты их хромосом. Почти одновременно был открыт парасексуальный процесс у грибов (Г. Понтекорво, Великобритания), что расширило возможности генетич. анализа грибов, не имеющих полового цикла размножения. Вскоре в генетич. исследования были вовлечены бактериофаги и др. вирусы (в частности, вирус табачной мозаики - ВТМ). Был открыт эффект переноса генетич. информации от одной бактериальной клетки к другой при посредстве бактериофага - генетич. трансдукция, что положило начало изучению генетических взаимоотношений в системе фаг - бактерия (Дж. Ледерберг, Н. Зиндер, США). Вслед за тем была обнаружена рекомбинация у фагов (А. Херши и М. Дельбрюк, США). Если использование бактерий в качестве объекта генетич. исследований резко повысило разрешающую способность генетич. анализа, то благодаря фагам удалось перейти к изучению явлений наследственности на молекулярном уровне. Большое значение имели исследования ВТМ (нем. генетики Г. Шустер и А. Гирер), позволившие вызвать генетич. эффект в опытах с чистой рибонуклеиновой к-той (РНК), к-рая сохраняла ннфекционность и при нанесении на листья табака вызывала в клетках образование полноценных частиц ВТМ. Исходя из общих принципов исследования генетпч. процессов у микроорганизмов, для каждой группы разработаны спец. методы изучения с учётом их особенностей . Генетич. механизмы у грибов и водорослей, сохранивших половой процесс, имеют ряд особенностей. Главная из них состоит в том, что продукты мейоза (споры) остаются соединёнными в определ. порядке, и после раздельного высева этих спор можно непосредственно изучать генотип каждого продукта мейоза. Этот метод, называемый тетрадным анализом, дополняет статистич. методы изучения процесса расщепления. Применение генетнч. анализа к организмам, у к-рых отсутствует половой процесс, стало возможным после открытия у них парасексуальных процессов, отличающихся большим разнообразием. Так, у несовершенных грибов при срастании гиф, принадлежащих двум генетически различным штаммам, происходит объединение и затем слияние двух гаплоидных ядер в одно днплоидное; в этой системе изредка возможен обмен генетич. материалом. Особенность полового процесса у бактерии состоит в том, что в клетку-реципиент передаётся, как правило, только часть генетич. материала из клетки-донора, в результате чего образуется частично диплоидная зигота (т. н. мерозигота). У бактерий известно неск. механизмов передачи генетнч. материала. Наиболее совершенная форма полового процесса у бактерий - конъюгация, детально изученная у кишечной палочки. Конъюгация происходит при непосредств. контакте между двумя клетками, если в одной из них присутствует специфич. половой фактор, или фактор скрещиваемости (фертильности, плодовитости). Половой фактор (см. Эписомы) содержит ДНК и может существовать в клетке либо в автономном, либо в интегрированном состоянии (включённым в геном клетки). В первом случае при конъюгации в клетку-реципиент переходит только половой фактор. Во втором случае половой фактор способствует направленному переносу генетич. материала из клетки-донора в клетку-реципиент. Как правило, при этом происходит передача только части генома донора и лишь крайне редко передается вся хромосома донора вместе с включённым в неё половым фактором. Между фрагментом донорной ДНК и ДНК реципиента может произойти обмен гомологичными генетич. участками - кроссинговер, приводящий к возникновению рекомбинантов, т. е. клеток с изменённым сочетанием признаков. Генетич. анализ рекомбинантов кишечной палочки позволил установить у неё существование одной группы сцепления, определить линейное расположение большого числа генов в её хромосоме и построить кольцевую генетич. карту (см. Генетические карты хромосом). Перенос генетич. материала при конъюгации - строго ориентированный процесс, при к-ром последовательность передачи генов (а значит, и вероятность их участия в кроссннговере) целиком зависит от расположения генов в хромосоме и точки интеграции (включения) полового фактора. При переходе полового фактора в автономное состояние гены, расположенные на хромосоме рядом с точкой интеграции, могут объединиться с половым фактором и в дальнейшем передаваться с ним как единое целое, превращая клетки-реципиенты в диплоиды по данному генетич. участку. Этот процесс переноса генов совместно с половым фактором, паз. сексдукцией, также может привести к возникновению рекомбинантов. Др. механизм возникновения рекомбинантов у бактерий - трансдукция - осуществляется при посредстве т. н. умеренных бактериофагов, к-рые способны к особому виду симбиоза с бактериями - лизогении. В лизогенных бактериях ДНК умеренного фага интегрирована с ДНК бактериальной клетки и ре-плицируется одновременно с ней. Такая скрытая форма присутствия фага (профаг) может сохраняться в течение мн. клеточных поколений, однако изредка профаг переходит в вегетативное состояние (т. е. начинает размножаться) и разрушает бактерию. При этом возможны захват небольшого фрагмента ДНК клетки-хозяина и последующий его перенос в др. клетку, в к-рой перенесённый участок генома может вступить в генетич. обмен с гомологичной областью клетки-реципиента. Обычно при трансдукцпи передаются гены, расположенные в непосредств. близости от места локализации профага в хромосоме бактерии. Однако нек-рые фаги осуществляют трансдукцию, при к-рой любой участок генома бактерии с равной вероятностью может быть перенесён в др. клетку. Иногда сам процесс лизогенизацни, т. е. включения умеренного фага в геном бактерии, может сопровождаться приобретением клеткой новых свойств (см. Лизогенная конверсия), напр, вирулентности. Ещё один тип полового процесса у бактерий, называемый трансформацией,- перенос генетич. материала без посредства полового фактора или умеренного бактериофага с последующим возникновением рекомбинантов (вследствие генетич. обмена между пооникшим в клетку фрагментом ДНК и ДНК клетки-реципиента). Особенности генетических механизмов у вирусов бактерий - бактериофагов - делают их весьма удобной моделью для изучения воспроизведения и функционирования генетич. материала. Они очень просто устроены, быстро размножаются и имеют очень короткий жизненный цикл; поэтому генетика бактериофагов, в особенности фагов Т2, Т 4 и /\., исследована весьма детально. Бактериофаги скрещивают, заражая бактерии смесью двух или неск. мутантов фага. В этом случае, кроме исходных фаговых частиц, появляются рекомбинанты с изменёнными сочетаниями признаков. С помощью рекомбинационного анализа удалось построить генетич. карты для ряда бактериофагов. Оказалось, что молекула ДНК фага является его хромосомой. Изучение тонкой структуры гена, проведённое на фаге Т4 (С. Бензер, США), показало существование большого числа участков внутри гена, способных изменяться (мутировать) с разной частотой под действием различных мутагенов. Генетика вирусов животных и растений в значит, мере основывается на успехах в области генетики бактериофагов, но из-за технич. трудностей ещё не получила достаточного развития. Возможность получения рекомбинантов была показана у ДНК-содержащих вирусов группы оспы - осповакцины (при смешанном заражении клеток различными представителями этой группы), у вируса герпеса (между различными вариантами этого вируса), а также между обезьяньим опухолеродным вирусом SV40 и различными представителями аденовирусов. У РНК-содержащих вирусов животных показана возможность получения рекомбинантов между мутантами вируса ящура и полиомиелита, а также между различными вариантами вируса гриппа. Последнее открытие имеет особое значение, т. к. показывает возможные пути изменчивости этого вируса в природе. Из вирусов растений лучше всего изучен вирус табачной мозаики (ВТМ). В частности, полностью расшифрована последовательность аминокислот в белке ВТМ; удалось установить характер аминокислотных замещений, возникающих в белках оболочки у разных мутантов ВТМ. Работы, выполненные на ВТМ, явились важным этапом в изучении как механизма мутагенеза, так и природы генетического кода. В связи с развитием новой отрасли народного хозяйства - микробиологической промышленности - возникла прикладная Г. м., наз. также селекцией микроорганизмов. В сферу исследований были вовлечены новые формы микроорганизмов: пенициллы (Penicillium chrysogenum), актиномицеты (Actinomyces streptomycini, Act. rimosus и др.), актинофаги. У пенициллов и аспергиллов открыт парасексуальный процесс, у актиномицетов изучен механизм рекомбинации, открыты генетич. рекомбинация у актинофагов, генетич. трансдукция у актиномицетов. Проведены обширные исследования индуцированной изменчивости количеств, признаков у актиномицетов. В Сов. Союзе в работе по селекции микроорганизмов применяются такие генетич. методы, как получение индуцированных мутаций, гибридизация и заражение актиномицетов актинофагами. В итоге выведены высокоактивные штаммы, позволившие во много раз увеличить производство антибиотиков, аминокислот, витаминов и др. биологически активных веществ. В связи с возрастающим значением Г. м. и необходимостью развития микробиол. пром-сти в 1968 в Москве был организован Всесоюзный н.-и. ин-т генетики и селекции пром. микроорганизмов Главмикробиопрома, ставший ведущим науч. центром в этой области. Проблемы генетики и селекции микроорганизмов разрабатываются и в др. науч. учреждениях Москвы (Ин-т эпидемиологии и микробиологии им. Н. Ф. Гамалеи АМН СССР, Ин-т общей генетики и Ин-т атомной энергии им. И. В. Курчатова АН СССР), Ленинграда (Физико-технич. ин-т им. А. Ф. Иоффе АН СССР, кафедра генетики ЛГУ), Киева, Еревана и др. Г.м. сыграла важную роль в развитии совр. генетики, дополнив ряд положений генетики высших организмов. Г. м., в свою очередь, стала основой для развития молекулярной генетики. Лит.: Xэйс У., Генетика бактерий и бактериофагов, пер. с англ., М., 1965; Гольдфарб Д. М., Введение в генетику бактерий, М., 1966; Захаров И. А. и Квитко К. В., Генетика микроорганизмов, Л., 1967; Алиханян С. И., Современная генетика, М., 1967; его же, Селекция промышленных микроорганизмов, М., 1968; Браун В., Генетика бактерий, пер. с англ., М., 1968; Генетические основы селекции микроорганизмов, М., 1969. С. И. Алиханян, Л. Н. Майсурян. ГЕНЕТИКА ПОВЕДЕНИЯ, область науки о поведении, основывающаяся на законах генетики и изучающая, в какой степени и каким образом различия в поведении определяются наследственными факторами. Осн. методы исследования Г. п. на экспериментальных животных - селекция в сочетании с инбридингом (близкородственное скрещивание), при помощи к-рых изучаются механизмы наследования форм поведения, на человеке - статистич. и генеалогич. анализ в сочетании с близнецовым и цитогенетич. методами. Зависимость поведения от наследств, факторов - генное управление и контроль поведения - исследуется на различных уровнях организации живого: в биоценозах, популяциях, сообществах, на уровне организма, а также на физиологическом (орган, ткань, клетка) и молекулярном уровнях. Исследования Г. п. имеют существ, значение для учения об индивидуальных различиях высшей нервной деятельности и выявления относит, роли врождённых и индивидуально приобретённых особенностей поведения, для объяснения роли генетически обусловленных особенностей поведения животных в популяции (для обществ, животных - в стаде, стае и т. п.), а также для создания экспериментальных моделей нервных болезней. Лит.: Крушинский Л. В., Генетика и феногенетика поведения животных, в кн.: Актуальные вопросы современной генетики, М., 1966; Гэйтс Дж., Молекулярная психобиология, пер. с англ., М., 1969; Fаlconer D. S., Introduction to quantitative genetics, N. Y., 1960; Fuller J. Thompson W. R., Behavior genetics, N.Y. - L., 1960. Л. Г. Романова. ГЕНЕТИКА РАСТЕНИЙ, раздел генетики, изучающий наследственность и изменчивость высших растений (генетич. исследования грибов и водорослей обычно относят к генетике микроорганизмов). Для генетич. изучения растений, кроме методов, к-рыми пользуются в др. областях генетики (в частности гибридологического анализа), применяют след, методы. С помощью моносо много анализа определяют роль каждой хромосомы в наследовании и развитии различных признаков растении. Этим методом (разработанным на дурмане) пользуются при изучении ряда аллополиплоидов (нек-рых пшениц, хлопчатника), а также диплоидов (ячменя). Большое значение в Г. р. приобретает экспериментальный мутагенез, к-рый даёт огромное разнообразие новых форм, используемых в селекции, и ценный материал для изучения генетики отд. видов растений. С помощью мутантов составляют генетические карты хромосом; на них исследуют действие изменённого гена (в гомо- и гетерозиготном состоянии) на развитие отд. признаков в разных условиях среды, на физиол. и биохим. особенности растений. Изучение мутантов способствует выяснению эволюции того или иного вида. К методам исследования эволюции растений относятся также гибридизация и анализ конъюгации хромосоме мейозе у гибридов (неродств. хромосомы не конъюгируют). Важный метод - искусственный ресннтез существующих видов путём гибридизации и последующего удвоения числа хромосом (см. Полиплоидия). Значит, роль в эволюции растений, в т. ч. мн. культурных (пшеницы, овса, хлопчатника, картофеля, плодовых и др.), играет аллополиплоидия. После открытия действия алкалоида колхицина, препятствующего расхождению удвоившихся хромосом к разным полюсам клетки, для получения новых, иногда очень ценных форм широко используется автополиплоидия. Сочетая методы отдалённой гибридизации и цитогенетики, изучают роль отд. хромосом (и их участков) в наследовании признаков и разрабатывают приёмы, позволяющие получать вставки участков хромосом диких растений, обусловливающие развитие ценных признаков (напр., устойчивости к ржавчине), в хромосомы культурных растений. Роль ядра и цитоплазмы в наследовании и развитии признаков исследуют, применяя отдалённую гибридизацию и анализируя природу мужской цитоплазматич. стерильности, используемой при получении гетерозисных форм. В Г. р. широко исследуются апомиксис и явление самонесовместимости, т. е. неспособности растений к самооплодотворению, а также генетич. особенности растений само- и перекрёстноопылителей, вегетативно и апомиктически размножающихся форм. В Г. р. всё больше проникают идеи и методы молекулярной биологии (гибридизация ДНК, ДНК - РНК, изучение изозимов и др.). Методы популяционной генетики и биометрии применяют в Г. р. для разграничения генотипич. и паратипич. элементов в общей фенотипич. изменчивости признаков, что усиливает эффективность искусств, отбора. Все эти методы используют для улучшения хозяйственно ценных свойств с.-х. растений: урожайности, устойчивости к неблагоприятным условиям среды, ряда биохим. и технологич. особенностей растения (или его зерна), особенностей развития (озимость, яровость, раннеспелость и т. д.). Из высших растений генетически наиболее изучены кукуруза, арабидопсис (растение сем. крестоцветных, растительная дрозофила - модельный объект генетич. исследований), горох, томаты, ячмень. У этих растений методами гибридизации установлена локализация генов и составлены карты хромосом. Интенсивно изучается цитогенетика мягкой пшеницы - сложного 42-хромосомного аллополиплоида, возникшего в процессе эволюции при естеств. гибридизации трёх разных злаков с последующим удвоением числа хромосом у гибридов. Вклад Г. р. в селекцию огромен. Это, напр., использование гетерозиса в селекции кукурузы на основе мужской стерильности; введение высокоурожайным гибридам и сортам кормового ячменя генов, обеспечивающих высокое содержание лизина в зерне; создание низкорослых неполегающих высокоурожайных сортов пшеницы с использованием генов карликовости (зелёная революция в Индии и др. странах); выведение урожайных и сахаристых триплоидных гибридов сахарной свёклы. Лит.: Вавилов Н. И., Избр. произв., т. 1, Л., 1967; Мичурин И. В., Соч., т. 1 - 4, М., 1948; Брюбейкер Дж. Л., Сельскохозяйственная генетика, пер. с англ., М., 1966; Эллиот Ф., Селекция растений и цитогенетика, пер. с англ. ,М.. 1961: Мюнтцинг А., Генетика, пер. с англ., М., 1967; Уильяме У., Генетические основы и селекция растений, пер. с англ., М., 1968; Цитогенетика пшеницы и ее гибридов, М., 1971; Генетические основы селекции растений, М., 1971. В. В. Хвостова. ГЕНЕТИКА ЧЕЛОВЕКА, отрасль генетики, тесно связанная с антропологией и медициной. Г. ч. условно подразделяют на антропогенетику, изучающую наследственность и изменчивость нормальных признаков человеческого организма, и генетику медицинскую, к-рая изучает его наследственную патологию (болезни, дефекты, уродства и др.). Г. ч. связана также с эволюционной теорией, т. к. исследует конкретные механизмы эволюции человека и его место в природе, с психологией, философией, социологией. Из направлений Г. ч. наиболее интенсивно развиваются цитогенетика, биохимич. генетика, иммуногенетика, генетика высшей нервной деятельности, физиологич. генетика. В Г. ч. вместо классич. гибридологического анализа применяют генеалогический метод, к-рый состоит в анализе распределения в семьях (точнее, в родословных) лиц, обладающих данным признаком (или аномалией) и не обладающих им, что раскрывает тип наследования, частоту и интенсивность проявления признака и т. д. При анализе семейных данных получают также цифры эмпирического риска, т. е. вероятность обладания признаком в зависимости от степени родства с его носителем. Генеалогич. методом уже показано, что более 1800 морфологич., биохимич. и др. признаков человека наследуется по законам Менделя (см. Генетика). Напр., тёмная окраска кожи и волос доминирует над светлой; пониженная активность или отсутствие нек-рых ферментов определяется рецессивными генами, а рост, вес, уровень интеллекта и ряд др. признаков - полимерными генами, т. е. системами из мн. генов. Мн. признаки и болезни человека, наследующиеся сцепленно с полом, обусловлены генами, локализованными в Х- или Y-хромосоме. Таких генов известно ок. 120. К ним относятся гены гемофилии А н В, недостаточности фермента глюкозо-6-фосфат-дегидрогеназы, цветовой слепоты и др. Др. метод Г. ч.- близнецовый метод (см. Близнецы). Однояйцевые близнецы (ОБ) развиваются из одной яйцеклетки, оплодотворённой одним спермием; поэтому набор генов (генотип) у ОБ идентичен. Разнояйцевые близнецы (РБ) развиваются из двух и более яйцеклеток, оплодотворённых разными спермиями; поэтому их генотипы различаются так же, как у братьев и сестёр (сибсов). Сравнение внутрипарных различий между ОБ и РБ позволяет судить об относит, значении наследственности и среды в определении свойств человеческого организма. В близнецовых исследованиях особенно важен показатель конкордантности, выражающий (в %) вероятность обладания данным признаком одним из членов пары ОБ или РБ, если его имеет другой член пары. Если признак детерминирован преим. наследств, факторами, то процент конкордантности намного выше у ОБ, чем у РБ. Напр., конкордантность по группам крови, к-рые детерминированы только генетически, у ОБ равна 100%. При шизофрении конкордантность у ОБ достигает 67%, в то время как у РБ - 12,1%; при врождённом слабоумии (олигофрении) - 94,5% и 42,6% соответственно. Подобные сравнения проведены в отношении ряда заболеваний. Т. о., исследования близнецов показывают, что вклад наследственности и среды в развитие самых разнообразных признаков различен и признаки развиваются в результате взаимодействия генотипа и внешней среды. Одни признаки обусловлены преим. генотипом, при формировании др. признаков генотип выступает в качестве предрасполагающего фактора (или фактора, лимитирующего норму реакции организма на действия внешней среды). Геном человека включает неск. миллионов генов, способных к тому же по-разному влиять на развитие признаков. В результате мутаций и перекомбинации генов возникает присущее человеку разнообразие по самым разным признакам. Гены человека мутируют каждый с частотой от 1 на 100 000 до 1 на 10 000 000 гамет на поколение. Распространение мутаций среди больших групп населения изучает популяционная Г. ч., позволяющая составить карты распространения генов, определяющих развитие нормальных признаков и наследственных болезней. Особый интерес для популяционной Г. ч. представляют изоляты - группы населения, в к-рых по к.-л. причинам (географич., экономич., социальным, религиозным и др.) браки заключаются чаще между членами группы. Это приводит к повышению частоты кровного родства вступающих в брак, а значит, и вероятности того, что рецессивные гены перейдут в гомозиготное состояние и проявятся, что особенно заметно при малочисленности изолята. Исследования в области Г. ч. продемонстрировали наличие естественного отбора в человеческих популяциях. Однако отбор у человека приобретает специфич. черты; он интенсивно действует только на эмбриональной стадии (т. н. самопроизвольные аборты - отражение такого отбора). Отбор в человеческом обществе осуществляется посредством дифференциальной брачности и плодовитости, т. е. в результате взаимодействия социальных и биологич. факторов. Мутационный процесс и отбор обусловливают огромное разнообразие (полиморфизм) по ряду признаков, присущее человеку, что делает его с биологич. точки зрения необычайно пластичным и приспособленным видом. Широкое использование в Г. ч. цитологич. методов способствовало развитию цитогенетики, где осн. объект исследования - хромосомы, т. е. структуры клеточного ядра, в к-рых локализованы гены. Установлено (1946), что хромосомный набор в клетках тела человека (соматических) состоит из 46 хромосом, причём женский пол определяется наличием двух Х-хромосом, а мужской - Х-хромосомы и Y-хромосомы. В зрелых половых клетках находится половинное (гаплоидное) число хромосом. Митоз, мейоз и оплодотворение поддерживают преемственность и постоянство хромосомного набора как в ряду клеточных поколений, так и в поколениях организмов. В результате нарушений указанных процессов могут возникать аномалии хромосомного набора с изменением числа и структуры хромосом, что приводит к возникновению т.н. хромосомных болезней, к-рые нередко выражаются в слабоумии, развитии тяжёлых врождённых уродств, аномалий половой дифференцировки или обусловливают самопроизвольные аборты. Успехи в развитии Г. ч. сделали возможными предупреждение и лечение наследственных заболеваний. Один из эффективных методов их предупреждения - медико-генетическое консультирование с предсказанием риска появления больного в потомстве лиц, страдающих данным заболеванием или имеющих больного родственника. Достижения биохимич. Г. ч. раскрыли первопричину (молекулярный механизм) мн. наследственно обусловленных дефектов, аномалий обмена веществ, что способствовало разработке методов экспресс-диагностики, позволяющих быстро и рано выявлять больных, и лечения мн. прежде неизлечимых наследств, болезней. Чаще всего лечение состоит во введении в организм веществ, не образующихся в нём вследствие генетич. дефекта, или в составлении специальных диет, из к-рых устранены вещества, оказывающие токсич. действие на организм в результате наследственно обусловленной неспособности к их расщеплению. Мн. генетич. дефекты исправляются с помощью своеврем. хирургич. вмешательства или педагогич. коррекции. Практич. мероприятия, направленные на поддержание наследств, здоровья человека, на охрану генофонда человечества, осуществляются через систему медико-генетических консультаций. Осн. цель медико-генетич. консультирования - информировать заинтересованных лиц о вероятности риска появления в потомстве больных. К медико-генетич. мероприятиям относится также пропаганда генетич. знаний среди населения, т. к. это способствует более ответственному подходу к деторождению. Медико-генетич. консультация воздерживается от мер принудительного или поощрительного характера в вопросах деторождения или вступления в брак, принимая на себя лишь функцию информации. Большое значение имеет система мер, направленных на создание наилучших условий для проявления положит, наследств, задатков и предотвращение вредных воздействий среды на наследственность человека. Г. ч. представляет собой естественнонаучную основу борьбы с расизмом, убедительно показывая, что расы - это формы адаптации человека к конкретным условиям среды (климатич. и иным), что они отличаются друг от друга не наличием хороших или плохих генов, а частотой распространения обычных генов, свойственных всем расам. Г. ч. показывает, что все расы равноценны (но не одинаковы) с биологич. точки зрения и обладают равными возможностями для развития, определяемого не генетич., а социально-историч. условиями. Констатация биологич. наследственных различий между отд. людьми или расами не может служить основанием для к.-л. выводов морального, юридического или социального порядка, ущемляющих права этих людей или рас (см. Геноцид, Сегрегация). Лит.: Ниль Дж. и Шэлл У., Наследственность человека, пер. с англ., М., 1958; Канаев И. И., Близнецы, М.- Л., 1959; Штерн К., Основы генетики человека, пер. с англ., М., 1965; Маккьюсик В., Генетика человека, пер. с англ., М., 1967; Биология человека, пер. с англ., М., 1968; Эфроимсон В. П., Введение в медицинскую генетику, 2 изд., М., 1968; Основы цитогенетики человека, [М., 1969]; Li Ching-chun, Human genetics, N. Y., 1961. К. Н. Гринберг, А. А. Прокофьева-Бельговская. ГЕНЕТИКО-АВТОМАТИЧЕСКИЕ ПРОЦЕССЫ, вероятностные процессы, определяющие изменение частоты разных аллелей в популяции. В больших, свободно скрещивающихся популяциях в отсутствие отбора и давления мутаций соотношение аллелей, независимо от их абсолютной исходной частоты, должно сохраняться во всех поколениях. Однако в реальных, ограниченных по численности популяциях частота генов не остаётся постоянной не только под давлением мутаций и отбора, но и в силу случайных отклонений. Детальный анализ Г.-а. п. был проведён сов. генетиками Н. П. Дубининым (1931), Н. П. Дубининым и Д. Д. Ромашовым (1932), англ. - Р. Фишером (1931) и амер. - С. Райтом (1931). Случайные колебания частоты аллелей в популяции связаны с тем, что распределение аллелей между гаметами и комбинирование гамет в зиготе - вероятностные процессы. Г.-а. п. оказывают несистематич. эффект, т. к. частота аллелей в разных поколениях может повышаться или понижаться. В малых популяциях или в популяциях, к-рые распадаются под действием изоляционных механизмов на отдельные подгруппы, может происходить чисто случайная стабилизация аллелей (гомозиготы) или их элиминация; в результате довольно быстро проявляются новые стабилизированные комбинации генов. Наиболее отчётливо Г.-а. п. проявляются при возникновении новых изолированных популяций. Напр., в секте меннонитов (Ланкастер, штат Пенсильвания, США), насчитывающей ок. 8000 человек, значителен процент карликов с многопалостью (13% меннонитов гетерозиготны по гену, к-рый в гомозиготном состоянии обусловливает появление таких карликов); это объясняется тем, что члены секты вступают в брак только между собой, а такая изоляция способствует появлению гомозиготных индивидуумов. В больших популяциях Г.-а. п. не могут обусловить такой стабилизации или элиминации аллелей, т. к. влияние этих процессов компенсируется за счёт разных факторов в последующих поколениях или в разных подразделениях популяции. Теория Г.-а. п. объяснила генетич. последствия изоляции, судьбу рецессивных мутаций на уровнях малых концентраций и эволюцию популяций по нейтральным признакам. Г.-а. п. объясняют мн. расовые различия человека, возникшие без действия отбора. Наряду с термином Г.-а. п. широко используется термин дрейф генов, предложенный С. Райтом. Сов. генетик С. С. Четвериков, подчёркивая роль вероятностно-статистич. закономерностей при дрейфе генов, предлагал назвать это явление генетико-стохастич. процессами. Лит.: Дубинин Н. П., Эволюция популяции и радиация, М., 1966, с. 421 - 33. Н. П. Дубинин, В. Н. Сойфер. ГЕНЕТИКОВ И СЕЛЕКЦИОНЕРОВ ОБЩЕСТВО Всесоюзное имени Н. И. Вавилова (ВОГИС), научно-общественная организация при АН СССР, объединяющая учёных и практиков СССР, работающих в области генетики и селекции. Создано в 1965. 30-31 мая 1966 в Москве состоялся учредительный съезд общества; был утверждён устав общества, избраны центральный совет (80 чел.) и президиум (23 чел.; находится в Москве). Президентом избран Б. Л. Астауров. К началу 1971 общество насчитывало 3670 чл., имело 26 отделений в столицах союзных республик, краевых, областных центрах и др. городах. Цели и задачи ВОГИС: активное участие в развитии всех отраслей генетики и селекции, повышение квалификации членов общества и реализация их исследований, популяризация и пропаганда новейших теоретич. и практич. достижений в области генетики и селекции, содействие преподаванию генетики и селекции в средней и высшей школе. В. Ф. Мирек. ГЕНЕТИЧЕСКАЯ ИНФОРМАЦИЯ, заложенная в наследственных структурах организмов (в хромосомах, цитоплазме, клеточных органеллах), получаемая от предков в виде совокупности генов информация о составе, строении и характере обмена составляющих организм веществ (прежде всего белков и нуклеиновых кислот) и связанных с ними функциях. У многоклеточных форм при половом размножении Г. и. передаётся из поколения в поколение через посредство половых клеток - гамет, единственная функция к-рых - передача и хранение Г. и. У микроорганизмов и вирусов имеются особые типы передачи Г. и. (см. Сексдукция, Трансдукция, Трансформация). Г. и. заключена преим. в хромосомах, где она зашифрована в определённой линейной последовательности нуклеотидов в молекулах дезоксирибонуклеиновой кислоты - ДНК (см. Генетический код). Г. и. реализуется в ходе онтогенеза - развития особи - передачей Г. и. от гена к признаку. Все клетки организма возникают в результате делений единственной исходной клетки - зиготы - и потому имеют один и тот же набор генов - потенциально одну и ту же Г. и. Специфичность клеток разных тканей определяется тем, что в них активны разные гены, т. е. реализуется не вся Г.и., а только её часть, необходимая для функционирования данной ткани. Ю. С. Дёмин. ГЕНЕТИЧЕСКИЕ КАРТЫ ХРОМОСОМ, схемы относительного расположения сцепленных между собой наследств, факторов - генов. Г. к. х. отображают реально существующий линейный порядок размещения генов в хромосомах (см. Цитологические карты хромосом) и важны как в теоретич. исследованиях, так и при проведении селекционной работы, т. к. позволяют сознательно подбирать пары признаков при скрещиваниях, а также предсказывать особенности наследования и проявления различных признаков у изучаемых организмов. Имея Г. к. х., можно по наследованию сигнального гена, тесно сцепленного с изучаемым, контролировать передачу потомству генов, обусловливающих развитие трудно анализируемых признаков; напр., ген, определяющий сморщенный эндосперм у кукурузы и находящийся в 9-й хромосоме, сцеплен с геном, определяющим пониженную жизнеспособность растения. Многочисленные факты отсутствия (вопреки Менделя законам) независимого распределения признаков у гибридов второго поколения были объяснены хромосомной теорией наследственности. Гены, расположенные в одной хромосоме, в большинстве случаев наследуются совместно и образуют одну группу сцепления, количество к-рых, т. о., соответствует у каждого организма гаплоидному числу хромосом (см. Гаплоид). Амер. генетик Т. X. Морган показал, однако, что сцепление генов, расположенных в одной хромосоме, у диплоидных организмов (см. Диплоид) не абсолютное; в нек-рых случаях перед образованием половых клеток между однотипными, или гомологичными, хромосомами происходит обмен соответств. участками; этот процесс носит назв. перекреста, или кроссинговера. Обмен участками хромосом (с находящимися в них генами) происходит с различной вероятностью, зависящей от расстояния между ними (чем дальше друг от друга гены, тем выше вероятность кроссинговера и, следовательно, рекомбинации). Генетич. анализ позволяет обнаружить перекрест только при различии гомологичных хромосом по составу генов, что при кроссинговере приводит к появлению новых генных комбинаций. Обычно расстояние между генами на Г. к. х. выражают как % кроссинговера (отношение числа мутантных особей, отличающихся от родителей иным сочетанием генов, к общему кол-ву изученных особей); единица этого расстояния - морганида - соответствует частоте кроссинговера в 1% . Г. к. х. составляют для каждой пары гомологичных хромосом. Группы сцепления нумеруют последовательно, номере их обнаружения. Кроме номера группы сцепления, указывают полные или сокращённые назв. мутантных генов, их расстояния в морганидах от одного из концов хромосомы, принятого за нулевую точку, а также место центромеры. Составить Г. к. х. можно только для объектов, у к-рых изучено большое число мутантных генов. Напр., у дрозофилы идентифицировано св. 500 генов, локализованных в её 4 группах сцепления, у кукурузы - ок. 400 генов, распределённых в 10 группах сцепления (рис. 1). У менее изученных объектов число обнаруженных групп сцепления меньше гаплоидного числа хромосом. Так, v домовой мыши выявлено ок. 200 генов, образующих 15 групп сцепления (на самом деле их 20); у кур изучено пока всего 8 из 39. У человека из ожидаемых 23 групп сцепления (23 пары хромосом) идентифицировано только 10, причём в каждой группе известно небольшое число генов; наиболее подробные карты составлены для половых хромосом. У бактерий, к-рые являются гаплоидными организмами, имеется одна, чаще всего непрерывная, кольцевая хромосома и все гены образуют одну группу сцепления (рис. 2). При переносе генетич. материала из клетки-донора в клетку-реципиент, напр, при конъюгации, кольцевая хромосома разрывается и образующаяся линейная структура переносится из одной бактериальной клетки в другую (у кишечной палочки в течение 110-120 мин). Искусственно прерывая процесс конъюгации, можно по возникшим типам рекомбииантов установить, какие гены успели перейти в клетку-реципиент. В этом состоит один из методов построения Г. к. х. бактерий, детально разработанных у ряда видов. Ещё более детализированы Г. к. х. нек-рых бактериофагов. См. также Генетика, Мутация. Рис. 1. Генетические карты 7-10-й хромосом кукурузы. Цифры по длине хромосом обозначают расстояние от конца хромосомы в морганидах; буквы - сокращённые названия признаков , определяемых соответствующими генами. Рис. 2. Генетическая карта хромосомы кишечной палочки (Eschenchia coli К 12). Цифры означают время (в мин), необходимое для переноса в клетку - реципиент генетических маркёров, контролирующих биосинтез ряда аминокислот, а также устойчивость к стрептомицину и к фагу Т6 эти цифры характеризуют расстояние между генами. Обозначения: ade - аденин; his - гистидин; try - триптофан; gal - галактоза; lac - лактоза; pro - пролин; leu - лейцин; tre - треонин; met - метионин; агд - аргинин; mt - маннит; xyl - ксилоза; mal - мальтоза; ser - серии; gly - глицин; sir иТ6, - устойчивость к стрептомицину или фагу Т6. Лит,.: Лобашев М. Е., Генетика, 2 изд., Л., 1967; Медведев Н. Н., Практическая генетика, 2 изд., М., 1968; Актуальные вопросы современной генетики. Сб. ст., М., 1966; Жакоб Ф., Вольман Э., Пол и генетика бактерий, пер. с англ., М., 1962; Бензер С., Тонкая структура гена, в сб.: Молекулярная генетика, пер. с англ., М., 1963; Xэйс У., Генетика бактерий и бактериофагов, пер. с англ., М., 1965; Рейвин А. У., Эволюция генетики, пер. с англ., М., 1967; Мюнтцинг А., Генетика, пер. с англ.,2 изд., М., 1967; Уотсон Дж., Молекулярная биология гена, пер. с англ., М., 1967. В. С. Андреев. ГЕНЕТИЧЕСКИЕ РЯДЫ, группы органич. соединений с одинаковым числом атомов углерода в молекуле, различающихся функциональными группами; см. Гомологические ряды. ГЕНЕТИЧЕСКИЙ АНАЛИЗ, совокупность методов изучения наследственных свойств организмов. Г. а. включает: 1) Гибридологический метод, изучающий законы наследственности, а также строение и поведение наследств, структур с помощью спец. видов скрещиваний (см. Гибридологический анализ). 2) Цитогенетический метод, развившийся на стыке генетики и цитологии. Главная его задача - установление связи между закономерностями наследования и строением и функциями хромосом (составление цитологических карт хромосом, геномный анализ и др.). 3) Молекулярно-генетический метод, получивший развитие в связи с новыми биохим. и физ.-хим. методами анализа наследственных структур. С его помощью изучается связь между молекулярным строением генов и синтезируемыми в соответствии с заложенной в них информацией белками. Лит.: Серебровский А. С., Генетический анализ, М., 1970 (библ.). Ю. С. Дёмин. ГЕНЕТИЧЕСКИЙ КОД, система зашифровки наследственной информации в молекулах нуклеиновых кислот, реализующаяся у животных, растений, бактерий и вирусов в виде последовательности нуклеотидов. В природных нуклеиновых кислотах - дезоксирибонуклеиновой (ДНК) и рибонуклеиновой (РНК) - встречаются 5 распространённых типов нуклеотидов (по 4 в каждой нуклеиновой к-те), различающихся по входящему в их состав азотистому основанию (см. Пуриновые основания, Пиримидиновые основания). В ДНК встречаются основания: аденин (А), гуанин (Г), цитозин (Ц), тимин (Т); в РНК вместо тимина присутствует урацил (У). Кроме них, в составе нуклеиновых к-т обнаружено ок. 20 редко встречающихся (т. н. неканонических, или минорных) оснований, а также необычных Сахаров. Т. к. кол-во кодирующих знаков Г. к. (4) и число разновидностей аминокислот в белке (20) не совпадают, кодовое число (т. е. кол-во нуклеотидов, кодирующих 1 аминокислоту) не может быть равно 1. Различных сочетаний по 2 нуклеотида возможно лишь 42 = 16, но этого также недостаточно для зашифровки всех аминокислот. Амер. учёный Г. Гамов предложил (1954) модель триплетного Г. к., т. е. такого, в к-ром 1 аминокислоту кодирует группа из трёх нуклеотидов, наз. кодоном. Число возможных триплетов равно 43 = 64, а это более чем втрое превышает число распространённых аминокислот, в связи с чем было высказано предположение, что каждой аминокислоте соответствует неск. кодонов (т. н. вырожденность кода). Было предложено много различных моделей Г. к., из к-рых серьёзного внимания заслуживали три модели (см. рис.): перекрывающийся код без запятых, неперекрывающийся код без запятых и код с запятыми. В 1961 Ф. Крик (Великобритания) с сотрудниками получил подтверждение гипотезы триплетного неперекрывающегося кода без запятых. Установлены след. осн. закономерности, касающиеся Г. к.:
Модели генетического кода: 1-й тип - перекрывающийся код без запятых; 2-й тип - неперекрывающийся код без запятых; 3-й тип - код с "промежутками", т. е. код с запятыми. 1) между последовательностью нуклеотидов и кодируемой последовательностью аминокислот существует линейное соответствие (колинеарность Г. к.); 2) считывание Г. к. начинается с определённой точки; 3) считывание идёт в одном направлении в пределах одного гена; 4) код является неперекрывающимся; 5) при считывании не бывает промежутков (код без запятых); 6) Г. к., как правило, является вырожденным, т. е. 1 аминокислоту кодируют 2 и более триплетов-синонимов (вырожденность Г. к. уменьшает вероятность того, что мутационная замена основания в триплете приведёт к ошибке); 7) кодовое число равно трём; 8) код в живой природе универсален (за нек-рыми исключениями). Универсальность Г. к. подтверждается экспериментами по синтезу белка in vitro. Если в бесклеточную систему, полученную из одного организма (напр., кишечной палочки), добавить нуклеиновокислотную матрицу, полученную из др. организма, далеко отстоящего от первого в эволюционном отношении (напр., проростков гороха), то в такой системе будет идти белковый синтез. Благодаря работам амер. генетиков М. Ниренберга, С. Очоа, X. Корана известен не только состав, но и порядок нуклеотидов во всех ко донах (см. табл., построенную по данным опытов с кишечной палочкой). Из 64 кодонов у бактерий и фагов 3 кодона - УАА, УАГ и УГА - не кодируют аминокислот; они служат сигналом к освобождению полипептидной цепи с рибосомы, т. е. сигнализируют о завершении синтеза полипептида. Их наз. терминирующими кодонами. Существуют также 3 сигнала о начале синтеза - это т. н. инициирующие кодоны - АУГ, ГУГ и УУГ, - к-рые, будучи включёнными в начале соответствующей информационной РНК (и-РНК), определяют включение формилметионина в первое положение синтезируемой полипептидной цепи. Приведённые данные справедливы для бактериальных систем; для высших организмов многое ещё не ясно. Так, кодон УГА у высших организмов может быть значащим; не совсем понятен также механизм инициации полипептида. Реализация Г. к. в клетке происходит в два этапа. Первый из них протекает в ядре; он носит назв. транскрипции и заключается в синтезе молекул и-РНК на соответствующих участках ДНК. При этом последовательность нуклеотидов ДНК "переписывается" в нуклеотидную последовательность РНК. Второй этап - трансляция - протекает в цитоплазме, на рибосомах; при этом последовательность нуклеотидов и-РНК переводится в последовательность аминокислот в белке; этот этап протекает при участии транспортной РНК (т-РНК) и соответствующих ферментов (см. Белки, раздел Биосинтез).
В начале цепи и-РНК данный кодон определяет начало синтеза полипептидной цепи и кодирует аминокислоту формилметионин. От готовых полиплоидных цепей формильная группа или вся аминокислота может быть отщеплена с помощью соответствующих ферментов. Лит.: Общая природа генетического кода для белков, в сб.: Молекулярная генетика, пер. с англ., М. 1963; Крик Ф., Генетический код (I), в кн.: Структура и функция клетки, пер. с англ., М., 1964, с. 9 - 23; Ниренберг М., Генетический код (II), там же, с. 24 - 41; Xэйс У., Генетика бактерий и бактериофагов, пер. с англ., М., 1965; Хартман Ф., Саскайнд З., Действие гена, пер. с англ., М., 1966; Бреслер С.Е., Введение в молекулярную биологию, 2 изд., М. - Л., 1966; Ингрэм В., Биосинтез макромолекул, пер. с англ., М., 1966; Лобашев М. Е., Генетика, 2 изд., Л., 1967; Уотсон Дж., Молекулярная биология гена, пер. с англ., М., 1967; Сойфер В. Н., Молекулярные механизмы мутагенеза, М., 1969; Дубинин Н. П., Общая генетика, М., 1970. Н. П. Дубинин, В. Н. Сойфер. ГЕНЕТИЧЕСКИЙ ПОЛИМОРФИЗМ, сосуществование в пределах популяции двух или нескольких различных наследственных форм, находящихся в динамич. равновесии в течение неск. и даже мн. поколений. Чаще всего Г. п. обусловливается либо варьирующими давлениями и векторами (направленностью) отбора в различных условиях (напр., в разные сезоны), либо повышенной относит, жизнеспособностью гетерозигот. Один из видов Г. п.- сбалансированный Г. п. - характеризуется постоянным оптимальным соотношением полиморфных форм, отклонение от к-рого оказывается неблагоприятным для вида, и автоматически регулируется (устанавливается оптимальное соотношение форм). В состоянии сбалансированного Г. п. у человека и животных находится большинство генов. Различают неск. форм Г. п., анализ к-рых позволяет определять действие отбора в природных популяциях. Лит.: Тимофеев-Ресовский Н. В., Свирежев Ю. М., О генетическом полиморфизме в популяциях, Генетика, 1967, № 10. ГЕНЕТИЧЕСКОЕ ДЕЙСТВИЕ ИЗЛУЧЕНИЙ, радиационный мутагенез, возникновение наследственных изменений (мутаций) при облучении организмов. Г. д. и.- важная часть биологического действия ионизирующих излучений, исследуемая радиационной генетикой. Первые стабильные радиорасы у дрожжей получены сов. биологами Г. А. Надсоном и Г. С. Филипповым (1925); данные о повышении частоты мутаций у дрозофилы при рентгеновском облучении опубликованы амер. генетиком Г. Меллером (1927). Мутагенный эффект вызывают все типы ионизирующих излучений, а также ультрафиолетовые лучи, если их действию подвергаются наследственные структуры любых организмов - от вирусов и бактерий до высокоорганизованных животных, включая человека; при этом у сложных организмов мутации могут возникать как в половых клетках - гаметах, так и в клетках тела - соматических. Облучение может вызывать все типы мутаций (генные, хромосомные, геномные и цитоплазматические). В определ. интервале доз частота мутаций возрастает пропорционально дозе облучения; при увеличении дозы выше нек-рого значения линейность кривых, описывающих зависимость частоты мутаций от дозы, нарушается. Вновь возникающие мутации являются обычно рецессивными (см. Рецессивность) и вредными. Повышение радиоактивного фона ведёт к накоплению в популяциях организмов, в т. ч. и человека, скрытых вредных мутаций. Важное практич. применение Г. д. и. - радиационная селекция, т. е. отбор хоз.-ценных мутаций, получаемых гл. обр. у культурных растений и пром. микроорганизмов в результате их облучения. Выведенные таким способом новые сорта овса, ячменя, гороха, арахиса, плодовых и декоративных культур и др. уже занимают большие посевные площади. Мн. высокопродуктивные пром. штаммы микроорганизмов - продуцентов антибиотиков, витаминов, аминокислот - также получены путём радиационного мутагенеза. Лит.: Итоги науки. Биологические науки, т. 3 - Ионизирующие излучения и наследственность, М., 1960; Астауров Б. Л., Функциональный принцип в оценке относительной значимости радиационных поражений ядра н цитоплазмы, в сб.: Первичные механизмы биологического действия ионизирующих излучений, М., 1963; Ли Д. Э., Действие радиации на живые клетки, пер. с англ., М., 1963; Алнханян С. И., Селекция промышленных микроорганизмов, М., 1968; Тимофеев-Ресовский Н. В., Иванов В. И., Корогодин В. И., Применение принципа попаданий в радиобиологии, М., 1968. Н. В. Тимофеев-Ресовский, В. И. Иванов. ГЕНЕТТА (Genetta), род хищных млекопитающих сем. виверровых. Длинное (до 55 см), приземистое и необычайно гибкое тело покрыто короткой, довольно грубой шерстью; окраска пятнистая; хвост не пушистый, до 50 см; у его основания расположены железы, выделяющие резко пахнущую жидкость-мускус. 6 видов; распространены гл. обр. в саваннах и тропич. лесах Африки. Обыкновенная Г. (G. genetta) широко распространена по всей Африке, встречается и в Юго-Зап. Европе (Испания, Франция), где населяет лесистые и безлесные горы и низменности, обитая преим. около водоёмов. По повадкам Г. напоминает хорьков. Питается мелкими зверьками, птицами и их яйцами, а также беспозвоночными. Иногда вредит птицеводству. Ведёт преим. ночной образ жизни. Г. легко приручаются; в Африке их иногда содержат дома для истребления крыс и мышей. Лит.: Mammals of the world, v. 2, Bait., 1964. ГЕНЗЕЛЬТ Адольф Львович (12.5.1814, Швабах, Бавария, - 10.10.1889, Вармбрунн, Силезия), русский пианист, педагог и композитор. Род. в нем. семье. В 1836 начал концертную деятельность в Берлине. С 1838 жил в Петербурге, занимаясь преим. преподаванием игры на фортепьяно (среди его учеников - В. В. Стасов, И. Ф. Нейлисов, Н. С. Зверев). В 1872-75 редактор муз. журнала "Нувеллист"; в 1887-88 профессор Петерб. консерватории. Игра Г. отличалась тонкой художеств. выразительностью, тщательной отделкой деталей и замечательным технич. мастерством. Фортепьянные произв. Г. (св. 40 опусов) мелодичны, изящны (особенно выделяются этюды, блестяще разработанные в пианистич. отношении), но подчас носят налёт салонной виртуозности. Г. принадлежит множество редакций, переложений и обработок для фортепьяно (в т. ч. обработки рус. нар. песен и произв. рус. композиторов). Автор инструктивно-педагогич. сочинения "На многолетнем опыте основанные правила преподавания фортепианной игры" (1868). Лит.: Ф. [Финдеизен Н.], Адольф Гензельт, Русская музыкальная газета, 1899, № 37; Алексеев А., Русские пианисты. Очерки и материалы по истории пианизма, в. 2, М.- Л., 1948; Музалевский В., Русская фортепианная музыка. Очерки и материалы по истории русской фортепианной культуры, Л.- М., 1949. ГЕНИАЛЬНОСТЬ, наивысшая степень проявления творческих сил человека. Термин Г. употребляется как для обозначения способности человека к творчеству, так и для оценки результатов его деятельности. Предполагая врождённую способность к продуктивной деятельности в той или иной области, гений, в отличие от таланта, представляет собой не просто высшую степень одарённости, а связан с созданием качественно новых творений, открытием ранее неизведанных путей творчества. Деятельность гения реализуется в определённом историч. контексте жизни человеческого общества, из к-рой гений черпает материал для своего творчества. В психологии творчества Г. изучается под углом зрения индивидуальных особенностей личности (её психич. склада, способностей и т. д.), а также разнообразных факторов, влияющих на творчество. С психологич. точки зрения гений не может рассматриваться как особый тип личности. Разнообразные попытки выделить к.-л. общие психологич. или психопатологич. черты Г. оказались неудовлетворительными: гениальные люди обнаруживают значительные индивидуальные различия с точки зрения одарённости, характера, культуры, интересов, навыков и т. д. Сам творческий процесс гения также принципиально не отличается с психологич. точки зрения от творческого процесса других одарённых людей. В ряде концепций Г., особенно начиная с Ч. Ломброзо, постулировалась связь между Г. и психич. расстройством, что, однако, не находит всеобщего подтверждения: дисгармония душевной жизни или предрасположенность к ней, свойственная многим гениальным людям, не является неизменным спутником Г. Историч. воззрения на природу Г. и её оценка связаны с общим пониманием творческого процесса (см. Творчество). Так, от античности идёт взгляд на Г. как род иррационального вдохновения, божественного наития (Платон, неоплатонизм). Начиная с эпохи Возрождения (Леонардо да Винчи, Дж. Вазари, Скалигер) получает распространение культ гения как творч. индивидуальности, достигающий своего апогея в период романтизма (предромантич. течение Бури и натиска в Германии, романтизм и вышедшие из него учения Т. Карлейля, Ф. Ницше с характерным для них противопоставлением гения и массы). В 18 в. складывается понятие гения в совр. смысле этого слова, к-рое у А. Шефтсбери становится одним из основных эстетич. понятий (гений творит подобно силе природы; его создания оригинальны в отличие от подражающего художника). И. Кант также подчёркивает оригинальность и естественность творчества гения: гений - это ...прирожденные задатки души..., через которые природа дает искусству правило (Соч., т. 5, М., 1966, с. 323). Ф. Шиллер раскрывает природу Г. через понятие наивности как инстинктивного следования безыскусств. природе и как способности к непредвзятому постижению мира. В 19-20 вв. получают развитие психологич. (в т. ч. психиатрические), социально-психологич., а также социология, исследования различных аспектов Г. и творчества. Лит.: Грузенберг С. О., Гений и творчество, Л., 1924; Жоли Г., Психология великих людей, СПБ, 1894; Оствальд В., Великие люди, [пер. с нем.], СПБ, 1910; Wо1f H.. Versuch einer Geschichte des Geniebegriffes, Hdlb., 1923; Genetic studies of genius, ed. L. M. Terman, 2 ed., v. 1-4, [Stanford], 1926-47; Kretschmer E., Geniale Menschen, 5Aufl., B., 1958. ГЕНИЙ (лат. genius), 1) в др.-рим. религии и мифологии добрый дух, сверхъестественное существо, охраняющее человека на протяжении всей его жизни. День рождения римлянина считался праздником его Г., к-рому приносились жертвы, совершались возлияния и курения. Римляне почитали также Г. - покровителей семьи, общины, города и всего римского народа. Культ Г. в Др. Риме - разновидность широко распространённого у многих народов в период разложения общинно-родовых отношений культа личных духов-покровителей. 2) Высшая степень творческой одарённости, а также человек, обладающий подобной одарённостью; см. Гениальность. ГЕНИСАРЕТСКОЕ ОЗЕРО, одно из названий Тивериадского озера, расположенного в Западной Азии. ГЕНИЧЕСК, город, центр Генического р-на Херсонской обл. УССР. Порт на Азовском м. Ж.-д. ст. 20 тыс. жит. (1970). Рыбоконсервный, винодельческий, арматурный, стройматериалов заводы. Мед. училище. Добыча соли. ГЕНИЧЕСКИЙ ПРОЛИВ, Тонкий пролив между Арабатской стрелкой и берегом, соединяет Азовское м. с зал. Сиваш. Дл. ок. 4 км, шир. 80-150 м. Глуб. 4,6 м. Течения зависят от ветров. На берегу - порт Геническ. ГЕНК (Genk), город в Бельгии, в пров. Лимбург. 56 тыс. жит. (1969). Добыча угля (копи Винтерслаг), металлургич., хим. промышленность. Автомобильный завод. ГЕНКИН Дмитрий Михайлович (19.9. 1884, Калуга,-24.1.1966, Москва), советский юрист, засл. деят. науки РСФСР (1945), доктор юридич. наук (1940). Чл. КПСС с 1964. Специалист в области гражд. права. С 1909 на научной и преподават. работе, в 1919 проф. Ин-та нар. х-ва им. Г. В. Плеханова (в 1919-21 ректор этого ин-та); проф. Всесоюзной академии внешней торговли, пред. Внешнеторговой арбитражной комиссии. Автор научных трудов, гл. обр. по теории гражд. права, правовым вопросам внешней торговли, правовому регулированию деятельности пром. предприятий. Награждён орденом Ленина, 3 др. орденами, а также медалями. Соч.: Законодательство о промысловой кооперации, М., 1930; Недействительность сделок, совершенных с целью, противной закону, Уч. зап. Всесоюзного ин-та юридических наук, 1947, в. 5; Право собственности в СССР, М., 1961; Правовые вопросы хозяйственного расчета государственных промышленных предприятий, М., 1966. ГЕННАДИ Григорий Николаевич [18(30). 3.1826, Петербург, - 26.2(9.3). 1880, там же], русский библиограф. Библиографич. деятельность начал в 1849. До кон. 50-х гг. занимался библиографией самых различных, не связанных между собой отраслей. С 60-х гг. работал в основном над "Справочным словарем о русских писателях и учёных". Крупнейшие труды Г.: Литература русской библиографии (1858) - систематич. аннотированный указатель рус. библиографич. лит-ры, и Справочный словарь о русских писателях и учёных, умерших в XVIII и XIX столетиях, и список русских книг с 1725 по 1825 [т. 1-3 (буквы А - Р), Берлин - М., 1876-1908, т. 4 остался в рукописи], содержащий сведения о рус. писателях и учёных, умерших до 1874, а также о рус. анонимных книгах, изданных в 1725-1825. Г. был одним из первых библиографов Пушкина, редактировал два изд. его соч., выпущенных Я. А. Исаковым (1859-60, 1869-71); в 1-м изд. Г. в качестве приложения опубликовал "Библиографический список всех произведений А. С. Пушкина..." (1860). Лит.: Ивасюк У. Г., Г. И. Геннади. Обзор жизни и трудов, М., 1913. ГЕННАДИЙ (г. рожд. неизв. - ум. 1505), русский церк. и политический деятель. Назначенный в 1484 новгородским архиепископом, Г. должен был стать проводником великокняж. политики в недавно присоединённом к Русскому централизованному гос-ву Новгороде. Но вскоре у Г. о вел. кн. Иваном III обнаружились противоречия из-за земельных владений архиепископской кафедры. Вместе с видным церковным деятелем Иосифом Волоцким Г. боролся против новгородско-моск. ереси (жидовствующих) и жестоко расправлялся с еретиками (см. Ереси). Публицистическая и переводческая деятельность лит. кружка, организованного Г., была связана с борьбой против еретиков. Обеспокоенный и смущённый малой грамотностью своих сподвижников и образованностью вольнодумцев, Г. выступал за создание училищ для духовенства и распространял церк.-полемич. и астрономич. лит-ру. При дворе Г. было написано Слово кратко в защиту церк. имуществ (первонач. название - Собрание на лихоимцев) и составлен первый в России полный библейский кодекс - Геннадиевская библия (1499). В 1504 Г. и его сподвижникам удалось добиться полного осуждения еретиков, но сам Г. по воле вел. князя был смещён с архиепископской кафедры. Умер в опале. Лит.: Слово кратко в защиту монастырских имуществ, Чтения в обществе истории и древностей российских, 1902, кн. 2, отд. 2; Собрание на лихоимцев - неизданный памятник русской публицистики конца XV в., Тр. Отдела древнерусской литературы, т. 21, М. - Л., 1965; Лурье Я. С.. Идеологическая борьба в русской публицистике конца XV - начала XVI вв., М.- Л., 1960. Я. С. Лурье. ГЕННЕГАУ (флам. Henegouwen, франц. Hainaut), ср.-век. графство, затем провинция ср.-век. Нидерландов, с 1830 - Бельгии; см. Эно. ГЕННЕП (van Gennep) Арнольд ван (23.4.1873, Людвигсбург, - 7.5.1957, Эперне), французский этнограф, фольклорист, исследователь первобытной религии. Президент общества франц. этнографов (1952-57), основатель ряда этнографич. изданий. Автор многочисл. трудов по общей этнографии и этнографии Франции; первым из франц. этнографов применил метод этнографич. картографирования. Соч.: Religions, moeurs et legendes. Essais d'ethnoaraphie et de linguistique, v. 1-5, P., 1908-14; Le folklore, P., 1924; Manuel de folklore francais contemporaine, pt. 1, 3, 4, P.. 1937-58. ГЕННИН, де Геннин Виллим Иванович (Георг Вильгельм) [11(21).10. 1676 - 12(23).4.1750], специалист и организатор горного и металлургического производства в России, генерал-лейтенант. Голландец по происхождению; с 1698 на рус. службе. Участвовал в Северной войне 1700-21. В 1713-22 был нач. горных заводов Олонецкого края. С 1722 нач. уральских горных заводов. При нём усовершенствованы старые заводы и выстроено 9 новых. После возвращения с Урала (1734) был управляющим Гл. арт. канцелярии, перестраивал тульские з-ды, заведовал сестрорецкими з-дами. Соч.: Описание уральских и сибирских заводов, М., 1937. Лит.: Павленко Н. И., Развитие металлургической промышленности России в первой половине XVIII в., М., 1953. ГЕНОВ (псевд. Цонев) Гаврил Димитров (1.2.1892, с. Живовци, Михайловградский округ, - 20.1.1934, Москва), деятель болг. рабочего движения. Сын крестьянина-бедняка. Работал сел. учителем, принимал участие (с 1912) в деятельности Видинской учительской орг-ции Болг. с.-д. партии (тесных социалистов). Во время Балканских войн 1912-13 и 1-й мировой войны 1914-18 - офицер; за антивоен. деятельность подвергался репрессиям. В 1920-23 секретарь Врачанского окружкома компартии. Во время Сентябрьского антифашистского восстания 1923 возглавлял вооружённую борьбу во Врачанском округе, был чл. Гл. революц. к-та. После поражения восстания эмигрировал в Югославию. В 1925-1926 чл. заграничного бюро ЦК БКП (тесных социалистов). С 1927 - в СССР, работал в Крестьянском интернационале и болг. секции ИККИ. Окончил междунар. Ленинскую школу. Редактировал перевод соч. В. И. Ленина на болг. язык. Соч.: Сентябрьское восстание в Болгарии 1923 года, М.- Л.,1934 (совм. с А. Владимировым). М. А. Бирман. ГЕНОГЕОГРАФИЯ, направление исследований в пограничной между генетикой и биогеографией области, сформулированное А. С. Серебровским в 1928- 1929. Осн. задача Г.- установление геогр. распространения и, по возможности, частот аллелей, определяющих осн. признаки и свойства в пределах всего или части ареала изучаемого вида организмов. Г. изучает также причины распространения аллелей. Проведение гено-геогр. работ возможно лишь у тех видов, у к-рых в экспериментально-генетич. исследованиях установлена связь между изучаемыми признаками и генами. Шире возможности т. н. феногеографин, изучающей геогр. распространение элементарных признаков в пределах ареала вида. Г. (и феногеография) имеет большое теоретич. значение в исследованиях по систематике и эволюции видов. Практич. и прикладное значение Г. имеет в установлении генофондов домашних животных и культурных растений как одной из основ породного и сортового районирования и селекции, а также в генетике человека и, особенно, в мед. генетике. Лит.: Серебровский А. С., Геногеография и генофонд сельскохозяйственных живвтных СССР, Научное слово, 1928, № 9; его же. Проблемы и метод геногеографии, в кн.: Труды Всесоюзного съезда по генетике, селекции, семеноводству и племенному животноводству, т. 2, Л., 1930. Н. В. Тимофеев-Ресовский. ГЕНОКОПИЯ, миметические гены, возникновение сходных фенотипич признаков под влиянием генов, расположенных в разных участках хромосомы или в разных хромосомах (т. н. мутантные аллели). Явление Г., установленное прежде всего на высших организмах, свидетельствует о сложном характере наследования мн. признаков. Биохим. природа Г. заключается в наличии в клетке неск. параллельных путей синтеза тех или иных её компонентов (напр., синтез тимидиловой к-ты в бактериальной клетке может осуществляться как из уридиловой, так и из цитидиловой к-т). Разные мутации, действие к-рых реализуется через один и тот же процесс или орган, могут с неодинаковой полнотой копировать друг друга по своему конечному эффекту; в свою очередь, их конечный эффект может имитироваться при действии различных внеш. факторов (см. Фенокопия). Явления Г. и фенокопии очень важны для понимания механизмов реализации наследственных (при Г.) и ненаследственных (при фенокопиях) аномалий и болезней у человека. В. Н Сойфер, В. П. Эфроимсон. ГЕНОМ [нем. Genom, англ, genom(e)], гаплоидный хромосомный набор; совокупность генов, локализованных в одиночном наборе хромосом данного организма. Термин предложен в 1920 нем. биологом Г. Винклером. Под Г. принято понимать совокупность генов, сосредоточенных в хромосомах без учёта наследственных детерминант, связанных со структурами цитоплазмы. В гаметах диплоидных организмов, а также в клетках гаплоидных организмов содержится один Г.; в соматич. клетках диплоидных организмов - два Г. С увеличением степени плоидности клеток растёт число Г. При оплодотворении происходит объединение Г. отцовских и материнских гамет. Как правило, Г., полученные от отцовской и материнской гамет, гомологичны. Гомология между всеми или неск. Г. отсутствует лишь у отдалённых гибридов. Под абс. гомологией двух Г. понимают совпадение линейного расположения генов в каждой хромосоме. Наличие такого совпадения обеспечивает возможность нормальной конъюгации хромосом в мейозе. Изменения числа хромосом (напр., полиплоидизация, увеличение числа или выпадение отдельных хромосом) наз. геномными мутациями. Организм, у к-рого неск. раз повторяется один и тот же Г., наз. автополиплоидом (см. Автополиплоидия). Организм, в к-ром объединены разные Г., наз. аллополиплоидом (см. Аллополиплоидия). Примером аллополиплоидов могут служить пшеницы. Гаплоидное число хромосом у твёрдой пшеницы - 14, у мягкой - 21, у однозернянки - 7. Путём гибридизации и изучения конъюгации хромосом в мейозе было выяснено, что один Г. из 7 хромосом имеется у всех пшениц (геном А); у твёрдой и мягкой - два общих Г. по 7 хромосом (А, В) и, наконец, у мягкой есть ещё особый геном D, также включающий 7 хромосом. Т. о., геномная формула однозернянки - АА, твёрдой - ААВВ, мягкой пшеницы, возникшей в процессе эволюции путём скрещивания трёх разных диких злаков и удвоения числа хромосом у гибридов,- AABBDD. В опытах сов. генетика Г. Д. Карпеченко были впервые совмещены в гибридном организме Г. редьки и капусты. Путём соответств. скрещиваний и цитологич. анализа можно установить происхождение отдельных Г. Напр., В. А. Рыбин от скрещивания алычи и тёрна получил (ресинтезировал) культурную сливу; т. о. было установлено, что Г. сливы включает Г. алычи и тёрна (см. Амфидиплоиды, Геномный анализ). Для понимания структуры и функционирования Г. большое значение имело установление строения молекул нуклеиновых кислот (ДНК и РНК), механизмов их репликации, способов записи и передачи генетич. информации (см. Генетический код). В. Н. Сойфер, В. В. Хвостова. ГЕНОМНЫЙ АНАЛИЗ, анализ происхождения различных геномов у полиплоидных форм посредством скрещиваний между предполагаемыми родительскими формами с последующей полиплоидизацией (см. Плоидность, Полиплоидия). Один из цитогенетич. методов; предложен япон. цитогенетиком X. Кихарой (1924). При подборе пар для скрещивания ориентируются на морфологич. признаки, общие для обоих партнёров и изучаемой аллополишюидной формы (см. Аллополиплоидия). После скрещивания и полиплоидизации проводят точный хромосомный анализ и анализ процесса мейоза. Конъюгация хромосом служит показателем родства геномов. Отсутствие конъюгации свидетельствует о принадлежности хромосом к разным геномам. ГЕНОНЕМА (от ген и греч. пета - нить), длинная белковая молекула-нить (или их пучок), к-рая, согласно модели, предложенной в 1928 сов. биологом Н. К. Кольцовым, представляет основу хромосомы и является носителем генетич. информации. Радикалы Г. рассматривались как гены, а атомные изменения в белковой молекуле - как причины мутаций. Эта модель после открытия роли дезоксирибонуклеиновой кислоты устарела, но выдвинутое Кольцовым при её построении предположение о способности хромосомы к самокопированию было подтверждено дальнейшими исследованиями. ГЕНОТИП (от ген и тип), совокупность всех генов, локализованных в хромосомах данного организма. В более широком смысле Г.- совокупность всех наследственных факторов организма - как ядерных (геном), так и неядерных, внехромосомных (т. е. цитоплазматических и пластидных наследственных факторов). Термин предложен датским биологом В. Иогансеном (1909). Г.- носитель наследственной информации, передаваемой от поколения к поколению. Он представляет собой систему, контролирующую развитие, строение и жизнедеятельность организма, т. е. совокупность всех признаков организма- его фенотип. Г. - единая система взаимодействующих генов, так что проявление каждого гена зависит от генотипической среды, в к-рой он находится. Напр., красная окраска цветков у нек-рых сортов душистого горошка возникает только при одновременном присутствии в Г. доминантных аллелей двух различных генов, тогда как порознь каждая из этих аллелей обусловливает белую окраску цветков (см. Комплементация). Взаимодействие Г. с комплексом факторов внутр. и внеш. среды организма обусловливает фенотипическое проявление признаков. Примером влияния среды на фенотипич. проявление Г. может служить окраска меха у кроликов т. н. гималайской линии: при одном и том же Г. эти кролики при выращивании на холоде имеют чёрный мех, при умеренной темп-ре - гималайскую окраску (белая с чёрными мордой, ушами, лапами и хвостом), при повыш. темп-ре - белый мех. Потомки этих трёх групп животных наследуют не какую-то одну неизменную окраску меха, а способность давать определ. окраску, различную в разных условиях среды. Поэтому в общем виде правильнее говорить, что Г. определяет наследование не конкретных признаков, а норму реакции организма на все возможные условия среды. На разных этапах развития особи в активном состоянии находятся то одни, то др. гены; поэтому Г. в онтогенезе функционирует как изменчивая подвижная система. Термин Г. иногда употребляют в более узком смысле для обозначения лишь группы генов или даже отдельных генов, наследование к-рых составляет предмет наблюдения. Напр., в расщепляющемся потомстве от моногибридного скрещивания АА X аа принято говорить о генотипах АА, Аа и аа, отвлекаясь от возможных различий между соответствующими особями (или группами особей) по др. генам. В.И.Иванов. ГЕНОТИПИЧЕСКАЯ СРЕДА, генетический фон, совокупность генов, оказывающих влияние на проявление в фенотипе (структурах и функциях организма) данного гена. Термин предложен сов. генетиком С. С. Четвериковым в 1926. В дальнейшем экспериментально установлено, что проявление каждого гена зависит от влияния др. генов. Это, в частности, доказано существованием генов-регуляторов. С углублением исследований клетки как целостной системы выясняется всё большее значение роли Г. с., прежде всего во взаимосвязанных процессах обмена веществ. ГЕНОФОНД (от ген и фонд), качественный состав и относительная численность разных форм (аллелей) различных генов в популяциях и населениях того или иного вида организмов. Термином Г. (введён рус. учёным А. С. Серебровским в 1928) обозначают аллельный состав популяции или всего населения вида, включая все варьирующие признаки и свойства вида или же ту или иную интересующую исследователя выборку из них. Рецессивный Г. - в основном укрытая от естеств. отбора совокупность рецессивных аллелей - при резком сокращении численности особей и изменившихся внеш. условиях обеспечивает быструю наследственную перестройку популяции. У генетически менее изученных видов можно определять т. и. фенофонд (фен - элементарный признак). Инвентаризация наследственно варьирующих признаков изучаемого вида с.-х. животных и растений совместно с определением частот различных аллелей имеет большое практич. значение. Изучение Г. человека имеет фундаментальное значение для генетики человека. Лит.: Серебровский А. С., Геногеография и генофонд сельскохозяйственных животных СССР, Научное слово, 1928, № 9. Н. В. Тимофеев-Ресовский. ГЕНОЦИД (от греч. genos - род, племя и лат. caedo - убиваю), истребление отдельных групп населения по расовым, национальным или религиозным мотивам, одно из тягчайших преступлений против человечества. Преступления Г. органически связаны с фашизмом и аналогичными реакционными теориями, пропагандирующими расовую и национальную ненависть и нетерпимость, господство т. н. высших рас над низшими и т. п. Преступления Г. совершались в массовых масштабах гитлеровцами во время 2-й мировой войны 1939-45 в оккупированных странах Европы, особенно против славянского и еврейского населения. Миллионы людей различных национальностей были уничтожены в фашистских лагерях смерти. Правящие круги ряда империалистич. гос-в грубо нарушают Конвенции 1948 и 1965, проводя политику Г. как внутри страны, так и на подвластных им территориях, используя эту политику для борьбы против национально-освободительного движения. Политика Г. и апартхейда стала гос. политикой в Южно-Африканской Республике, в Родезии. Наказуемость Г. установлена уставами междунар. военных трибуналов (Нюрнбергского и Токийского), а также спец. междунар. конвенцией О предупреждении преступления геноцида и наказании за него (одобрена Генеральной Ассамблеей ООН 9 дек. 1948). Согласно конвенции, под Г. понимаются действия, совершаемые с намерением уничтожить, полностью или частично, к.-л. национальную, этническую, расовую или религиозную группу как таковую, а именно: убийство членов такой группы, причинение им серьёзных телесных повреждений или умственного расстройства; предумышленное создание условий, к-рые рассчитаны на полное или частичное физич. уничтожение таких групп, принятие мер, рассчитанных на предотвращение деторождения в их среде, насильственная передача детей из одной человеческой группы в другую. Конвенция предусматривает также наказуемость заговора с целью совершения Г., подстрекательства, покушения и соучастия в Г. Конвенция предусматривает предупреждение ц наказание преступлений физич. и биол. Г. При выработке конвенции представитель СССР настаивал на запрещении также национально-культурного Г., к-рый выражается в мероприятиях и действиях, направленных против пользования национальным языком и против национальной культуры к.-л. группы населения, однако империалистич. державы отказались принять это предложение, а также распространить действие конвенции на колонии, в к-рых преступления Г. носят массовый характер. В 1965 ООН принята конвенция о ликвидации всех форм расовой дискриминации, осуждающая расизм, одной из форм которого является Г. В Советском Союзе и других социалистич. гос-вах всякое ограничение прав или установление к.-л. привилегий граждан в зависимости от их расовой, национальной или религиозной принадлежности, а также проповедь расовой или нац. исключительности или ненависти и пренебрежения запрещены и караются законом. ГЕНРИ (Henry) Джозеф (17.12.1797, Олбани, - 13.5.1878, Вашингтон), американский физик. С 1832 проф. Прин-стонского колледжа, с 1846 секретарь и директор Смитсоновского ин-та, с 1868 президент Нац. АН США; первый президент Философского об-ва в Вашингтоне (с 1871). Осн. труды по электротехнике. В 1828 впервые построил электромагниты большой силы, применив многослойные обмотки из изолированной проволоки. Г. открыл явление самоиндукции (1832) и колебательный характер разряда конденсатора (1842). Его именем названа единица индуктивности в системе СИ - генри. Г. принадлежат также работы по метеорологии. Соч.: Scientific writings, v. 1 - 2, Wash., 1886. Лит.: Лебедев В. И., Изобретение Джозефа Генри, Вестник связи, 1946, № 8. ГЕНРИ (О. Henry) (1862-1910), американский писатель; см. О. Генри. ГЕНРИ (Henry) Уильям (12.12.1774, Манчестер, - 2.9.1836, Пендлбери), английский химик. Доктор медицины Эдинбургского ун-та (1807). Открыл зависимость растворимости газов в воде от температуры и давления (см. Генри закон). ГЕНРИ, единица индуктивности и взаимной индуктивности в Международной системе единиц и МКСА системе единиц. Названа в честь амер. учёного Дж. Генри. Сокращённое обозначение: русское гн, междунар. Н. 1 генри равен индуктивности электрич. контура, возбуждающего магнитный поток в 1 вебер при силе постоянного тока в нём 1 ампер. Г. может быть также определён как индуктивность электрич. цепи, в к-рой возникает эдс самоиндукции в 1 вольт при равномерном изменении тока в этой цепи со скоростью 1 ампер в 1 сек. На практике часто пользуются дольными единицами: миллигенри (10-3 гн) и микрогенри (10-6 гн). Генри на метр - единица абсолютной магнитной проницаемости, равная абс. магнитной проницаемости среды, в к-рой при напряжённости магнитного поля 1 а/м создаётся магнитная индукция 1 тл. Г. Д. Бурдун. ГЕНРИ ЗАКОН, положение, согласно к-рому при постоянной темп-ре растворимость газа в данной жидкости (выраженная весовой концентрацией его) прямо пропорциональна давлению этого газа над раствором. Г. з. описан У. Генри в 1803; он хорошо соблюдается только для идеальных растворов и применим лишь в области невысоких давлений, приобретая при этом характер предельного закона. О растворимости газовых смесей см. Дальтона законы. В. А. Киреев. ГЕНРИЕТТЫ ОСТРОВ, один из о-вов архипелага Де-Лонга в Восточно-Сибирском м. Пл. 12 км2. Сложен гл. обр. песчаниками. Вые. 315 м. Покрыт ледниками. ГЕНРИХ (Henry). В Англии: Г. I [1068, Селби (?), - 1.12.1135, Лион-ла-Форе], король с 1100. Младший сын короля Вильгельма I Завоевателя. Незаконно захватил власть, склонив на свою сторону англ. баронов обещанием гарантировать соблюдение их феод, прав и привилегий. Опираясь на мелкое рыцарство, растущие города и свободное крестьянство, Г. I значительно укрепил гос. аппарат. При нём оформился центр, орган управления - королевская курия. Г. I вступил в конфликт с римским папой по вопросу о праве назначения англ, епископов. По соглашению 1107 король сохранил право на получение феод, присяги от англ, прелатов при их вступлении во владение церк. землями. Г. II (5.3.1133, Ле-Ман, - 6.7.1189, Шинон), король с 1154, первый из династии Плантагенетов. Под властью Г. II наряду с англ, владениями находились обширные владения во Франции. Став королём после долгой феодальной смуты (1135-53), Г. II восстановил мир и добился роспуска баронских наёмных отрядов. Опираясь на рыцарство, горожан и верхушку свободного крестьянства, проводил политику дальнейшего укрепления централизованного феод, гос-ва в Англии. Судебная реформа Г. II значительно расширила компетенцию королев, судов за счёт феод, курий и ввела в них расследование через присяжных; отныне любой свободный мог за плату обратиться в королев, суд. Попытка Г. II с помощью т. н. Кларендонских постановлений (1164) подчинить королев, власти церк. суды не увенчалась успехом. В результате воен. реформы широкое распространение получила замена рыцарской службы ден. платежом, т. н. щитовыми деньгами; кроме того, все свободные были обязаны иметь оружие, чтобы при необходимости служить в войске короля. При Г. II были заложены основы всей суд.-адм. системы англ. феод, гос-ва: королев, курия разделилась на высший королев, суд и казначейство, начало складываться англ, общегос. феод, право. При всей прогрессивности реформ Г. II они носили чисто феод, характер. Проводя резкую грань между свободными и крепостными, они юридически закрепляли бесправие последних. При Г. II в 1169 началось завоевание Ирландии. Лит.: Boussard J., Le gouvernement d'Henry II, Plantagenet, P., 1956. E. В. Гутнова. Г. III (1.10.1207, Уинчестер, - 16.11. 1272, Лондон), король (с 1216) из династии Плантагенетов, сын Иоанна Безземельного. Пытался управлять страной, опираясь на иностр. феодалов-авантюристов и союз с рим. курией. Недовольство баронов этой политикой в кон. 50-х гг. нашло поддержку у мелкого рыцарства, горожан и зажиточного крестьянства. В 1258 Г. III был вынужден утвердить Оксфордские провизии, установившие в стране режим баронской олигархии, а затем Вестминстерские провизии, неск. ограничивавшие произвол баронов. Отказ Г. III от соблюдения •Оксфордских провизии привёл к гражд. войне (1263-67). Симон де Монфор в 1264 разбил королевские войска и взял в плен Г. III. После разгрома сторонниками короля войск оппозиции и гибели Монфора (авг. 1265) Г. III в 1266 был полностью восстановлен в своих правах. Однако возросшая роль рыцарства и городов вынудила Г. III и баронов установить практику созыва парламента. Лит.: Гутнова Е. В., Возникновение английского парламента, М., 1960. Е. В. Гутнова. Г. IV [апр. 1366(?), замок Болингброк, графство Линкольншир, - 20.3.1413, Лондон], король с 1399, основатель династии Ланкастеров. В 1397 поддержал гос. переворот Ричарда II, но вскоре был изгнан. В 1399 высадился в Англии и возглавил мятеж магнатов Сев. Англии против Ричарда II. После отречения последнего был провозглашён королём. Правление Г. IV прошло в борьбе с мятежами крупных феодалов, восстаниями в Уэльсе, набегами шотландцев. Г. IV искусно разъединял своих противников, стремился опираться на мелкое дворянство и зажиточных горожан и заручиться поддержкой парламента, права к-рого были значительно расширены. Жестоко подавлял демократич. движения; в 1401 провёл статут, направленный против лоллардов. Ю. Р. Ульянов. Г. V[16.9(?). 1387, Монмут, - 31.8.1422, Буа-де-Венсенн], король (с 1413) из династии Ланкастеров, сын Генриха IV. Стремился примириться с феод, знатью, возвратив ей владения, конфискованные Генрихом IV. В 1414 подавил восстание лоллардов. Возобновив Столетнюю войну, нанёс французам в 1415 решит, поражение при Азенкуре и вскоре захватил С. Франции с Парижем. Был признан наследником франц. короля Карла VI и регентом Франции. Ю. Р. Ульянов. Г. VI (6. 12.1421, Виндзор,-21.5.1471, Лондон), король (1422-61) из династииЛанкастеров, сын Генриха V. Вступил на престол 9-месячным ребёнком. Слабовольный и подверженный всё более продолжительным припадкам безумия, был игрушкой в руках часто сменявшихся регентов и фаворитов. В 1461, в ходе Алой и Белой розы войны, был низложен королём Эдуардом IV Йорком. После врем, реставрации (3.10.1470 - 11.4.1471) Г. VI был вторично низложен и убит в Тауэре. Г. VII (28.1.1457, Пембрук, - 21.4. 1509, Ричмонд), король с 1485, основатель династии Тюдоров. Вступил на престол как кандидат Ланкастеров после победы над Ричардом III в битве при Босворте (22 авг. 1485), знаменовавшей завершение войны Алой и Белой розы. Провёл ряд мероприятий, ограничивавших могущество феод, знати [конфискация зем. владений, роспуск воен. дружин, создание чрезвычайных судов для расследования дел о заговорах (Звёздная палата, с. 1487)]. В интересах нарождавшейся буржуазии способствовал развитию судоходства и внешней торговли. В правление Г. VII были заложены основы абсолютизма. В. Ф. Семёнов. Г. VIII (28.6.1491, Гринвич, - 28.1. 1547, Лондон), король с 1509, 2-й из династии Тюдоров; один из ярких представителей англ, абсолютизма. В своём правлении опирался на узкий круг фаворитов (Т. Уолси, а позднее Т. Кромвель и Т. Кранмер). В правление Г. VIII была проведена Реформация, к-рую он рассматривал как важное средство укрепления абсолютизма и королев, казны. Поводом к Реформации послужил отказ папы утвердить развод Г. VIII с Екатериной Арагонской и его женитьбу на Анне Болейн. В 1534 после разрыва с папой был провозглашён парламентом главой английской (англиканской) церкви, сохранившей католич. обряды. Т. Мор (лорд-канцлер с 1529), выступивший против Реформации, был казнён (1535). В 1536 и 1539 была проведена секуляризация монастырей, земли к-рых перешли в руки нового дворянства. Сопротивление этой политике, особенно на С., жестоко подавлялось (см. Благодатное паломничество). Громадные расходы двора, войны с Францией и Шотландией привели в конце царствования Г. VIII к полному расстройству финансов. В связи с усилившейся в результате секуляризации экспроприацией крестьянства издал статуты против бродяг и нищих (см. Кровавое законодательство против экспроприированных). Хотя политика Г. VIII отвечала в нек-рой степени интересам нового дворянства и растущей буржуазии, его классовой опорой было феод, дворянство (попытки Г. VIII сохранить старую феод. структуру землевладения в эпоху начавшегося агр. переворота отразились, в частности, в его мероприятиях по ограничению огораживаний). Лит.: Масkie J. D.. The earlier Tudors. 1485-1558, Oxf., 1952; Scar i sbrick J. J., Henry VIII,L., 1969. В. Ф. Семёнов. ГЕНРИХ (Heinrich). В королевстве Германия и Священной Римской империи. Наиболее значительны: Г. I (ок. 876 - 2.7.936, Мемлебен, Саксония), король с 919, основатель Саксонской династии. Саксонский герцог из рода Людольфингов. От вынужденной политики уступок усилившимся герцогам Г. I постепенно переходил к их подчинению. Вновь присоединил (925) к Герм, королевству отпавшую ранее Лотарингию. Для борьбы с набегами венгров выстроил и укрепил ряд бургов и создал сильную конницу; одержал победу над венграми при Риаде на р. Унструт (933). Воен. походами 928-929 начал захват земель полабских славян. Политика Г. I подготовила значит, усиление королев, власти при его сыне и преемнике Оттоне I. Лит.: Bartmuss H. J.. Die Geburt des ersten deutschen Staates, В., 1966. Г. III (28.10.1017 - 5.10.1056), король с 1039, император с 1046, из Фоанконской династии; сын Конрада II. Опирался на министериалов и рыцарство. Во время похода в Италию (1046-47) низложил соперничавших пап, после чего неоднократно назначал кандидатов на папский престол. Однако покровительство Г. III церк. клюнийской реформе подготовило почву для последующего усиления папской власти. Поставил Чехию и Венгрию в зависимость от империи; вынудил к подчинению герцога Лотарингского. Г. IV (11.11.1050-7.8.1106, Льеж), король с 1056, император с 1084, сын Генриха III. В период малолетства Г. IV в Германии усилились князья, расхищавшие владения короны. Мероприятия Г. IV по укреплению позиций королев, власти в Саксонии вызвали Саксонское восстание 1073-75, с трудом им подавленное. Стремление сохранить за собой право церк. инвеституры в Германии и Сев. Италии привело Г. IV (в 1076) к столкновению с папой Григорием VII; разговелась длит, борьба за инвеституру. Отлучённый папой от церкви и низложенный, Г. IV был вынужден под давлением князей пойти на покаяние к папе в Каноссу (янв. 1077). Вновь отлучённый от церкви (1080), в 1084 овладел Римом и был коронован своим ставленником антипапой Климентом III, но отступил перед союзниками Григория VII - норманнами Юж. Италии. Объединение пап с нем. князьями, неудачный 3-й поход в Италию (1090-97), восстания против Г. IV его сыновей и его пленение не заставили изворотливого и энергичного Г. IV признать себя побеждённым. Ему удалось бежать из плена, но во время подготовки новой войны с сыном он умер. В целом попытка Г. IV упрочить королев, власть (путём укрепления королев, домена, опоры на министериалитет и отчасти на города) окончилась полным крахом в условиях роста раздробленности Германии. Лит.: Schmeidler R., Kaiser Heinrich IV, Lpz., 1927; Stern L. und Gericke H.. Deutschland von der Feudalepoche von der Mitte des 11. Jahrhunderts bis zur Mitte des 13. Jahrhunderts, В., 1964. М. Л. Абрамсон. Г. V (8.1.1081 - 23.5.1125, Утрехт), король с 1106, император с 1111, сын Генриха IV. При жизни отца сблизился с его противниками - папой и нем. князьями. После смерти Генриха IV возобновил борьбу с папством за инвеституру, окончившуюся компромиссным Вормсским конкордатом (1122). Со смертью Г. V прекратилась Франконская династия. F.VI (1165, Нимвеген, - 28.9.1197, Мессина), король с 1190, император с 1191 из династии Штауфенов. Сын Фридриха I Барбароссы. Благодаря женитьбе на наследнице сицилийского короля Констанции (1186) присоединил к владениям Штауфенов Сицилийское королевство (утвердился там лишь в 1194). Г. VII (ок. 1275 - 24.8.1313, Буонконвенто, близ Сиены), король с 1308, император с 1312, первый из династии Люксембургов. Добился в 1310 передачи своему сыну Иоанну чеш. престола. В 1310 вторгся в Италию, безуспешно пытаясь силой вновь подчинить её империи. ГЕНРИХ (Henri). Во Франции. Наиболее значительны: Г. II (31.3.1519, Сен-Жермен-ан-Ле, - 10.7.1559, Париж), король с 1547. Из династии Валуа. Был женат (с 1533) на Екатерине Медичи. Находился под влиянием коннетабля Анн де Монморанси, особенно в области внешней политики. Жестоко преследовал гугенотов, учредив для суда над ними в 1547 Огненную палату, в 1559 издал эдикт, требовавший для еретиков смертной казни. В союзе с немецкими протестантскими князьями вёл борьбу против имп. Карла V; в 1552 занял епископства Мец, Туль, Верден. В 1558 отвоевал у англичан Кале. В 1559 подписал Като-Камбрезийский мир, завершивший Итальянские войны. Был смертельно ранен на турнире. Лит.: Noell H., Henri П..., Р., 1944. Г. III (19.9.1551, Фонтенбло, - 2.8.1589, Сен-Клу), король с 1574, последний представитель династии Валуа. Был избран в 1573 на польск. престол, но, узнав о смерти своего брата франц. короля Карла IX, тайно бежал из Польши, чтобы занять франц. престол. Г. III правил в разгар Религиозных войн. Боролся как с гугенотами, к-рых возглавлял Генрих Наваррский, так и с Гизами, вождями Католической лиги, претендовавшими на наследование престола в связи с бездетностью Г. III. В мае 1588 в день баррикад (восстание в Париже, организованное Парижской лигой - объединением, игравшим решающую роль в Католич. лиге 1585-96) Г. III бежал в Шартр. В том же году после убийства герцога Гиза и его брата кардинала Лотарингского, совершённого по приказу Г. III, демократич. крыло Парижской лиги низложило Г. III. Последний пошёл на соглашение с Генрихом Наваррским, и они совместно осадили Париж. При осаде Г. III был убит подосланным лигой монахом Жаком Клеманом. Соч.: Lettres d'Henri III, t. 1 - 2, P., 1959-65. Лит.: Erlanger Ph., Henri III, P., [1935]: Champion P., Henri III, [v.1-2], P., [1943 - 51]. А. Д. Люблинская. Г. IV (13. 12. 1553, По, Беарн, - 14.5.1610, Париж), король с 1589 (фактически с 1594), первый из династии Бурбонов; король Наварры (Генрих Наваррский) с 1562. Во время Религиозных войн - глава гугенотов. В 1593 принял католичество и вступил в Париж. Нантским эдиктом 1598 предоставил гугенотам свободу вероисповедания и многие привилегии. Политика Г. IV способствовала укреплению абсолютизма. Ген. штаты перестали созываться; компетентность провинц. собраний была ограничена; возросла роль бюрократич. аппарата, чиновники добились официального закрепления права наследования и продажи должностей (см. Полетта). Г. IV отнял у губернаторов значит, долю гражданской власти, усилив полномочия ген. наместников и начинавших играть известную роль интендантов провинций. В области экономики проводил политику протекционизма, поощрял развитие франц. мануфактур; ввёл покровительств. таможенный тариф (1599); при нём проводились работы по улучшению дорог, строительству каналов и т. п. В 1604 началась колонизация французами Канады. Правительством Г. IV были отменены недоимки с крестьян за прошлые годы, снижена талья, но одновременно были увеличены косвенные налоги (на соль, вино и др.). Г. IV играл гл. роль в организации антигабсбургской оппозиции, вёл подготовку к войне с Габсбургами. Был убит католиком-фанатиком Равальяком. Генрих IV. Гравюра X. Голциуса. Лит.: Мосина З., Абсолютизм в политике Генриха IV, Историк-марксист, 1938, кн. 2; Пыхтеев Б., Мероприятия Генриха IV по развитию сельского хозяйства во Франции. Уч. зап. Московского государственного педагогического ин-та. 1940, т. 26; De Vaissiere P., Henri IV, P., [1928]; Ritter R., Henri IV, luimeme. L'Homme, P., [1944]. А. Д. Люблинская. ГЕНРИХ ЛАТВИЙСКИЙ (Heinrich von Lettland) (1187-1259), автор Хроники Ливонии (см. Ливонские хроники), написанной в 1224-27 на латинском яз. Онемеченный католич. священник. Прибыл в Ригу в 1205. С 1208 священник латышского прихода в Паппендорфе. Участник походов нем. крестоносцев в Ливонию. Восхвалял захватнич. политику рыцарей. В Хронике подробно описывается агрессия нем. феодалов в Ливонии и Эстонии с кон. 12 в. по 1227. Соч.: Heinrici Chronicon Livoniac. 2 Aufl., Hannoverae, 1955; в рус. пер. - Хроника Ливонии, М. -Л., 1938. Х. Х. Круус. ГЕНРИХ ЛЕВ (Heinrich der Lowe) (1129-6.8.1195, Брауншвейг), герцог Саксонии (в 1142-80) и Баварии (в 1156-80) из рода Вельфов. Вместе с Альбрехтом Медведем возглавил крестовый поход против славян 1147, окончившийся неудачей. В результате последующих походов (с 1160) захватил почти всю терр. бодричей и стал владельцем огромной терр. к В. от Эльбы. Истребляя и оттесняя славян к востоку, Г. Л. переселял на захваченные земли нем. колонистов. Усиление Г. Л. вызвало резкий конфликт между ним и имп. Фридрихом I Барбароссой. Последний, используя отказ Г. Л. от участия в походе императора в Италию, организовал суд. процесс над Г. Л. (1180). Г. Л. был лишён большинства владений (в его руках остались только Брауншвейг и Люнебург). ГЕНРИХ МОРЕПЛАВАТЕЛЬ (Dom Enrique о Nayegador) (4.3. 1394, Порту,- 13.11.1460, Сагрищ), португальский принц, организатор морских экспедиций к о-вам центр, части Атлантич. ок. и берегам Африки (за что в 19 в. получил прозвище Мореплаватель, хотя сам не плавал). Г. М. на средства Ордена Христа (главой к-рого он стал) основал в Сагрише (Португалия) обсерваторию и мореходную школу, способствовал развитию португ. кораблестроения (преобладающей стала каравелла). Океанские экспедиции Г. В. Кабрала, А. Кадамосто и др. открыли о-ва Азорские (1432-35), Зелёный мыс, pp. Сенегал и Гамбия, о-ва Бижагош (1434-57) и о-ва Зелёного Мыса (1456). По инициативе Г. М. начался вывоз афр. рабов в Португалию; капитаны Н. Триштан, Д. Диаш, А. Фернандиш и др., в поисках золота и охотясь за рабами, в 1434-60 обследовали и нанесли на карту ок. 3500 км зап.-афр. побережья от Зап. Сахары до Гвинейского зал. Экспедиции Г. М. положили начало португ. экспансии в Африку. Лит.: Магидович И. П., Очерки по истории географических открытий, М., 1967: Sanсеau E., Henry, the Navigator..., N. Y., 1947. ГЕНРИХ НАВАРРСКИЙ, король Наварры с 1562, франц. король Генрих IV. ГЕНСЛЕ (польск. gesle), 1) общее наименование старинных польских струнных инструментов. 2) Г. подгалянские, или злубцоки (ziobcoki), старинный польский народный смычковый инструмент. Корпус деревянный, долблёный грушевидный (как у болг. гадулки) или клеёный лодкообразный, струн 3-4, строй квинтовый. При игре инструмент опирали о левую сторону груди ниже плеча. Звук Г. - резкий, пронзительный. ГЕНТ, Ган (флам. Gent, франц. Gand), город в Бельгии, на р. Шельда при впадении в неё р. Лис, каналами связан с портами Остенде и Тернёзен на Северном м. Адм. ц. провинции Вост. Фландрия. 229,7 тыс. жит. (1969, с пригородами). Первое упоминание о Г. в источниках относится к 7 в. С 11-12 вв. Г. один из центров суконного произ-ва европ. значения. Непосредственно подчинённый власти фландрских графов, Г. в 12-13 вв. добился значит, самостоятельности в делах внутр. гор. управления и стал одним из гл. центров социально-политич. борьбы во Фландрии. Население Г. участвовало во Фландрском восстании 1323-28, здесь происходило восстание, возглавленное с 1338 Я. Артевелде, Гентское восстание 1539-40. Г. - один из центров революц. движения во время Нидерл. бурж. революции 16 в. В 16 в. в связи с общим упадком цехового ремесла Г. потерял значение одного из важнейших экономич. центров Европы; в 19 в. стал вновь экономически возрождаться. Г.- крупный пром. центр и трансп. узел. Второй по значению порт и гл. текст, центр страны. Преим. хл.-бум. и льняное произ-во. Металлургия, текст, и электро-технич. машиностроение, нефтепереработка, хим., бум., пищ. (муком., пивовар. и др.) пром-сть. Старинное произ-во кружев. Ввоз хлопка, льна, угля, нефти и др. Г.-центр фламандской культуры. Ун-т (с 1817). Музеи: археологич., изящных иск-в. Гент. Площадь Синт-Бафсплейн. На переднем плане - городская башня (1183-1339) с суконными рядами (1426 - 41,), за ними церковь Синт-Никласкерк (13- 17 вв.). Ср.-век. планировка, обилие архит. памятников начиная с романской эпохи (замок графов Фландрских, 1180-1200, - овальный в плане, с мощными стенами и башнями; хлебный склад, ок. 1200), старинная застройка площадей и набережных придают центру Г. вид города-музея. Особенно богат Г. готич. сооружениями (собор св. Бавона, ок. 1228-1559, с Гентским алтарём братьев ван Эйк; городская башня выс. 91 м, 1183-1339; ратуша, 1518-35, арх. Д. де Вагемакере, Р. Келдерманс; Дом свободных корабельщиков, 1530-31; укреплённый мост Работ, 1489; многочисл. церкви, монастырские постройки, жилые дома). Среди позднейших сооружений - Дворец юстиции (1836-46, арх. Л. Руландт), библиотека ун-та (1935-40, арх. X. ван де Велде; илл. см. т. 3, вклейка кс. 185), новые пром. р-ны. Лит.: Пиренн А., Средневековые города Бельгии, пер. с франц., М., 1937; D'Hоndt J., Keyser p. de, Gent, [Antw., 1947]; Fris V., Histoire de Gand, Brux., 1913; Keyser P. de, 1000-jarig Gent, Gent, 1949. ГЕНТЕКС (англ. Gentex - Generalized Teletype Exchange Service), международная телеграфная сеть общего пользования, оборудованная автоматич. коммутационными телеграфными станциями прямых соединений. Входящая в эту сеть телеграфная станция Г. в Москве соединена международными каналами связи с аналогичными станциями ряда европейских социалистич. стран. Др. телеграфные станции в Сов. Союзе получают соединение со станциями Г. набором их номеров через моек, станцию. ГЕНТ-ОСТЕНДЕ КАНАЛ, в Бельгии, соединяет р. Шельда у г. Гент с Северным м. у порта Остенде. Состоит из двух частей: канала Гент - Брюгге (прорыт в 1751) длиной в 47 км и канала Брюгге - Остенде (1622) длиной в 20,5 км. Глубина 3,1 м. Осн. грузы: уголь, стройматериалы, с.-х. продукция, хим. изделия, текст, сырьё. ГЕНТСКИЙ ДОГОВОР 1814, договор между Англией и США, завершивший англо-американскую- войну 1812-14. Подписан 24 дек. 1814 в Генте. Предусматривал взаимное возвращение захваченных территорий, прекращение воен. действий против индейцев, принятие решит, мер к прекращению работорговли. Восстанавливая старые границы, договор не разрешил спорных вопросов, вызвавших войну. За Г. д. последовало заключение торг, конвенции 1814, соглашения о демилитаризации Великих озёр 1817, конвенции 1818 о рыболовстве в Сев. Атлантике, о сев. границе США и т. н. совместном владении Орегоном. ГЕНТСКОЕ ВОССТАНИЕ 1539-40, было непосредственно вызвано попыткой исп. пр-ва Карла V насильственно собрать налог, на к-рый Гент не дал согласия. Но причины восстания лежали глубже (кризис цехового сукноделия, тяжёлые для трудящихся масс последствия процесса первоначального накопления капитала). Начало открытому восстанию положил захват власти в городе цехами (19 авг. 1539), его кульминацией был период 2 сент.- 3 нояб., характеризующийся политич. преобладанием гор. плебса и окрестной бедноты (крезеры). Восстание распространилось и на соседние города. Восставшие казнили, арестовали или изгнали наиболее ненавистных членов гор. олигархии, конфисковали их имущество. После попытки (хотя и неудавшейся) патрициата и богатого бюргерства организовать переворот восстание пошло на убыль. 14 февр. 1540 Карл V с войском вступил в Гент. Участники восстания подверглись репрессиям. Гент был обложен контрибуцией, лишён всех вольностей. Лит.: Чистозвонов А. Н., Гентское восстание 1539-1540 гг., М., 1957. А. Н. Чистозвонов. ГЕНТСКОЕ УМИРОТВОРЕНИЕ, соглашение, заключённое в г. Гент 8 нояб. 1576 в ходе Нидерландской буржуазной революции 16 века между сев. провинциями Нидерландов, восставшими в 1572, и юж. провинциями, на к-рые восстание распространилось в сент. 1576. Предусматривало совместную борьбу против исп. войск в Нидерландах, амнистию участникам восстания, сохранение католицизма на Ю. и кальвинизма на С. страны и др. Владения католич. церкви и власть Филиппа II Испанского формально оставались неприкосновенными. С созданием революц. Утрехтской унии (1579) Г. у. фактически утратило силу. ГЕНТСМАН, правильнее Хантсмен (Huntsman) Бенджамин (4. 6. 1704, Линкольншир, - 1776, Аттерклифф), автор тигельного способа получения литой стали (около 1740). Родился в Англии в немецкой семье. Тигельный процесс был известен, по-видимому, ещё в древности в странах Азии (Индия и др.), но технология его держалась в секрете, в Европе её не знали. Г. переплавлял куски сварочного железа и чугуна (выплавленного на древесном угле) в присутствии нек-рого количества флюса. Долгое время тигельный процесс был единственным способом получения литой стали. Лит.: Beck L., Die Geschichte des Eisens in technischer und kulturgeschichtlicher Beziehung, Abt. 3, Braunschweig, 1897. ГЕНТ-ТЕРНЁЗЕН КАНАЛ, в Бельгии и Нидерландах. Г.- Т. к. соединяет р. Шельда у г. Гент с её устьем у порта Тернёзен. Прорыт в 1827, реконструирован в 1911. Дл. 34 км (на терр. Бельгии 19,5 км). Ширина 50 м, глуб. 8,75 м. Осн. грузы: уголь, руды, строит, камень, химич. продукция, с.-х. продукты, текст, сырьё. ГЕНУЭЗСКАЯ КОНФЕРЕНЦИЯ 1922, международная конференция по экономическим и финансовым вопросам. Проходила в Генуе (Италия) с 10 апр. по 19 мая при участии представителей 29 государств - Австрии, Албании, Бельгии, Болгарии, Великобритании, Венгрии, Германии, Греции, Дании, Исландии, Испании, Италии, Латвии, Литвы, Люксембурга, Нидерландов, Норвегии, Польши, Португалии, РСФСР, Румынии, Королевства сербов, хорватов и словенцев, Финляндии, Франции, Чехословакии, Швейцарии, Швеции, Эстонии, Японии, а также 5 брит, доминионов - Австралии, Индии, Канады, Новой Зеландии и Юж.-Афр. Союза. Работой делегации РСФСР руководил В. И. Ленин, назначенный её председателем. Заместителем председателя делегации был Г. В. Чичерин, к-рый в Генуе, куда Ленин не выезжал, пользовался всеми правами председателя. Делегация РСФСР (в её состав входили также Л. Б. Красин, М. М. Литвинов, В. В. Воровский, Я. Э. Рудзутак, А. А. Иоффе, X. Г. Раковский, Н. И. Нариманов, Б. Мдивани, А. Бекзадян, А. Г. Шляпников) представляла на Г. к. интересы не только РСФСР, но и всех др. сов. республик (Азербайджанской, Армянской, Белорусской, Бухарской, Грузинской, Украинской, Хорезмской), а также интересы Дальневосточной Республики. США, отказавшиеся участвовать в работе Г. к. (нота гос. секретаря Ч. Юза от 8 марта 1922), были представлены на ней наблюдателем - амер. послом в Италии Р. Чайлдом. Среди представителей капиталистич. гос-в на Г. к. наиболее активную роль играли Д. Ллойд Джордж, Дж. Н. Керзон (Великобритания); К. Вирт, В. Ратенау (Германия); Л. Факта (Италия); Ж. Барту, К. Баррер (Франция). Решение о созыве Г. к. было принято по инициативе Великобритании на совещании Верх, совета Антанты в Канне (Франция) 6 янв. 1922 (см. Каннская конференция 1922). Офиц. целью Г. к. было изыскание мер к экономич. восстановлению Центр, и Вост. Европы. Но главным вопросом, стоявшим на конференции, был, по существу, вопрос об отношениях между Сов. государством и капиталистич. миром после провала попыток свержения Сов. власти путём воен. интервенции. Капиталистич. страны, в первую очередь Великобритания, в поисках преодоления послевоен. экономич. трудностей пытались вернуть на мировой рынок Сов. Россию (чтобы, пользуясь её временной экономической слабостью, широко эксплуатировать её ресурсы), а также Германию и бывших союзников Германии, потерпевших поражение в 1-й мировой войне 1914-18. Сов. правительство, заинтересованное в нормализации экономич. и политич. отношений с капиталистич. гос-вами, согласилось принять участие в работе Г. к. (8 янв. 1922). На конференции, однако, ведущую роль играли те представители капиталистич. стран, к-рые вместо делового обсуждения реальных путей установления экономии, связей с Сов. государством пытались с помощью дипломатич. нажима добиться от Сов. правительства экономич. и политич. уступок, ведущих к реставрации капитализма в России; они рассчитывали заставить Сов. государство признать все долги царского и Врем, пр-в, вернуть иностр. капиталистам национализированные Советской властью предприятия или возместить стоимость этих предприятий, ликвидировать монополию внеш. торговли и т. д. Сов. делегация по указанию В. И. Ленина отвергла эти требования и, в свою очередь, выдвинула контрпретензии о возмещении Сов. государству убытков, причинённых иностр. интервенцией и блокадой (если довоен. и воен. долги России были равны 18,5 млрд. золотых руб., то убытки Сов. государства в результате иностр. интервенции и блокады составляли 39 млрд. золотых руб.). Вместе с тем, желая найти почву для соглашения и восстановления экономич. связей с капиталистич. гос-вами, сов. делегация на Г. к. 20 апр. 1922 заявила, что Сов. правительство готово признать довоен. долги и преимущественное право за бывшими собственниками получать в концессию или аренду ранее принадлежавшее им имущество, при условии признания Сов. государства де-юре, оказания ему финанс. помощи и аннулирования воен. долгов и процентов по ним. Огромное значение имело внесённое сов. делегацией на первом же пленарном заседании Г. к. (10 апр. 1922) предложение о всеобщем разоружении. Однако как вопрос о разоружении, так и вопросы урегулирования взаимных финанс.-экономич. претензий по вине капиталистич. гос-в не были разрешены на Г. к. Обсуждение финанс.-экономич. вопросов было продолжено на Гаагской конференции 1922. В ходе Г. к. сов. дипломатии, использовавшей противоречия в империалистич. лагере, удалось прорвать единый фронт империалистич. гос-в, пытавшихся добиться дипломатич. изоляции Сов. государства, и заключить с Германией Рапалльский договор 1922 (см. в ст. Советско-германские договоры и соглашения). Публ.: Генуэзская конференция 1922. Материалы Генуэзской конференции (Подготовка, отчеты заседаний, работы комиссий, дипломатическая переписка и пр.), М., 1922; Документы внешней политики СССР, т. 5, М., 1961. Лит.: Ленин В. И., Проект директивы заместителю председателя и всем членам генуэзской делегации, Поли. собр. соч., 5 изд., т. 44; его же, О международном и внутреннем положении Советской Республики. Речь на заседании коммунистической фракции Всероссийского съезда металлистов 6 марта 1922 г., там же, т. 45; его же, Политический отчет ЦК РКП(б) 27 марта [на XI съезде РКП(б) 27 марта - 2 апреля 1922], там же; его же, Проект постановления ВЦИК по отчёту делегации на Генуэзской конференции, там же; Любимов Н. Н., Эрлих А. Н., Генуэзская конференция (Воспоминания участников), М., 1963; Рубинштейн Н. Л., Внешняя политика Советского государства в 1921 - 1925 гг., М., 1953; История дипломатии, 2 изд., т. 3, М.,1965, с. 249-304. И. И. Минц. ГЕНУЭЗСКАЯ ШКОЛА, одна из местных школ живописи в Италии 17 в. Сложилась к концу 1-й трети 17 в. под влиянием как традиций итальянской живописи 2-й половины 16 в., так и работавших в Генуе фламандских живописцев П. П. Рубенса и А. ван Дейка. Сочная живописность и чувственность фламандского искусства сказываются уже в ранних жанровых картинах Б. Строцци - ведущего художника Г. ш. старшего поколения. Генуэзские художники выполняли также парадные арнстократич. портреты, религ. композиции, декоративные росписи. Наиболее своеобразным созданием Г. ш. был пасторальный жанр, в к-ром большое внимание уделялось изображению пейзажа, домашней утвари и животных. Ведущими мастерами этого жанра были Дж. Б. Кастильоне и его последователь А. М. Вассалло, работавший в сер. 17 в. Тогда же работал и В. Кастелло: необычайно свободная, динамичная манера письма придаёт его композициям на религ. темы взволнованно-романтич., а порой и остродраматич. оттенок. Характерная для Кастелло живопись мазка и пятна получила распространение в Генуе и др. художеств, центрах Сев. Италии. Дж. Б. Кастильоне. Пастораль (фавн и пастушка). Музей изобразительных искусств им. А. С. Пушкина Москва. В. Кастелло. Мадонна с младенцем Христом и маленьким Иоанном. Музей изобразительных искусств им. А. С. Пушкина. Москва. Своеобразна генуэзская архит. школа, сложившаяся к сер. 16 в.; её расцвет связан с творчеством Г. Алесси, его учеников (Р. Лураго и др.) и последователей. Характерные черты генуэзской архитектуры наиболее ярко проявились в структуре домов генуэзской знати, сочетавших торжеств. пышность городского дворца с интимностью сельской усадьбы (см. Генуя). Лит.: Виппер Б. Р., Проблема реализма в итальянской живописи XVII - XVIII веков, М., 1966, с. 105 - 15; Всеобщая история архитектуры, т. 5, М., 1967, с. 259 - 63; Dе1оgu G., I pittori minori liguri, lombardi e piemontesi..., del seicento e del settecento, Venezia, 1931, p. 7 - 73. В. Э. Маркова. ГЕНУЭЗСКИЕ КОЛОНИИ В СЕВЕРНОМ ПРИЧЕРНОМОРЬЕ, укреплённые торг, центры генуэзских купцов в 13-15 вв. Генуэзцы в 1266 добились от ставленника Золотой орды в Крыму Мангу-хана передачи им во владение Кафы (совр. Феодосия), ставшей позже центром их колоний. В 1357 генуэзцы захватили Чембало (Балаклаву), в 1365 - Солдайю (Судак). Возникли новые колонии генуэзцев: Боспоро (на месте совр. Керчи), Тана (в устье Дона). В колониях жили греки, итальянцы, армяне, татары, русские и др. К кон. 14 в. они стали играть решающую роль в черноморской торговле. Генуэзские купцы вели обширную посредническую торговлю. Они продавали зерно, соль, кожи, меха, воск, мёд, лес, рыбу, икру из причерноморских р-нов, сукна - из Италии и Германии, масло и вино - из Греции, пряности, драгоценные камни, мускус - из стран Азии, слоновую кость - из Африки и мн. др. Большое место занимала торговля пленниками, выкупленными у тат. ханов и тур. султанов. Торг, операции генуэзских купцов производились также в рус. землях. Выходцы из генуэзских колоний - фряги - жили в Москве, где в 14-15 вв. существовала корпорация купцов -сурожан, специализировавшихся на торговле с Г. к. в С. П. Генуэзские колонии были хорошо укреплены, в крепостях имелись гарнизоны. Генуэзцы поддерживали союзнич. отношения с монголотат. ханами, к-рые формально являлись верховными владетелями терр. колоний, но предоставляли им полное самоуправление, сохраняя власть лишь над подданными ханов. В 1380 генуэзская пехота участвовала на стороне Мамая в Куликовской битве. Тем не менее Г. к. в С. П. Неоднократно подвергались нападениям и разорению со стороны таг. ханов (1299, 1308, 1344-1347, 1396-97 и др.). Торговая деятельность генуэзских купцов сочеталась с грабежом и эксплуатацией местного населения. Внутри самих колоний развивались глубокое социальное расслоение и острые нац.-религ. противоречия. Наиболее крупной колонией была Кафа, являвшаяся развитым центром ремесла. Во главе управления Кафой стояло богатое купечество, подчинившее и закабалившее массу бедноты. В 1433 в Чембало было крупное восстание местного населения против генуэзцев. В 1454 в Кафе произошло большое восстание гор. бедноты; дальнейшее обострение классовой борьбы привело к новым восстаниям в Кафе в 1456, 1463, 1471, 1472 и 1475. Углубление социальных и нац.-религ. противоречий предопределило упадок Г. к. в С. П. в 15 в. После падения Византийской империи (1453) междунар. положение колоний ухудшилось. В 1475 Г. к. в С. П. были захвачены и разгромлены Турцией и её вассалом Крымским ханством. Судак. Крепость 1345. Сохранились остатки крепостных стен, башен и дворцов в Кафе, Чембало и Солдайе, построенных невольниками под руководством итал. архитекторов. Крепость и консульский замок в Солдайе (14 в.) являются замечат. образцом итал. зодчества; там же уцелели остатки фресковых росписей. В 1951-52 в Кафе велись археологич. раскопки, давшие ценный материал для изучения истории города, его ремесла и торговли. Генуэзская башня в Балаклаве. 14 в. Лит.: Зевакин Е. С. и Пенчко Н. А.. Очерки по истории генуэзских колоний на Западном Кавказе вXIII и XVвв., в сб.: Исторические записки, т. 3, [М.], 1938 (библ.); их же, Из истории социальных отношений в генуэзских колониях Северного Причерноморья в XV в., там же, т. 7, [М.], 1940; Секиринский С., Очерки истории Сурожа XI - XV вв., Симферополь, 1955. А. М. Сахаров. ГЕНУЭЗСКИЙ ЗАЛИВ (Golfo di Genova), залив Лигурийского м. у сев.-зап. берегов Италии. Дл. 30 км; шир. у входа 96 км. Глуб. 1000-1500 м. Берега крутые и скалистые. Солёность 36,5o/00. Приливы полусуточные, их величина 0,3 м. Крупные порты - Генуя и Савона. ГЕНУЯ (Genova), город в Сев. Италии, на берегу Генуэзского зал. Лигурийского моря. Гл. город пров. Генуя и обл. Лигурия. 842,8 тыс. жит. (1969). Прижатый горами (Лигурийскими Апеннинами) к морю, город вытянулся более чем на 30 км вдоль узкой прибрежной полосы (Итальянской Ривьеры). Г. поглощает ближайшие к ней небольшие города (пром. пригороды к 3. от Г. - Корнильяно-Лигуре, Сампьердарена, Пельи, Вольтри, Сестри-Поненте; пром. города в горных долинах к С. от Г. - Больцането, Ривароло, Понтедечимо; курортные города к В. от города) и вместе с ними образует крупный урбанизированный р-н Большая Генуя. В пригороде Стальено - знаменитое кладбище (Кампо Санто). Г. - один из важнейших портов Средиземноморья, через к-рый осуществляются основные внешние связи пром. р-нов Севера страны, для к-рых он служит ближайшим выходом на внешний рынок. На Г. приходится св. 1/5 грузооборота всех итал. портов (45,4 млн. т в 1967, в т. ч. нефтяные грузы - 27,4 млн. т). Во ввозе преобладают нефть, уголь, металлолом, хлопок, лес, зерно, в вывозе - готовые пром. изделия. Г.- крупный пассажирский порт, узел ж.-д., автомоб. и воздушных сообщений. Г. - пром. и торг, центр. В Большой Г. размещается преим. тяжёлая пром-сть, отличающаяся высокой степенью концентрации произ-ва и господством нескольких крупных монополий (Финсидер, Финмеканика, Ансальдо и др.). Здесь сосредоточены крупнейшие в Италии судоверфи, произ-во самолётов, авиационных и судовых моторов, турбин, котлов, локомотивов, тракторов, электрооборудования; военная пром-сть, точная механика, металлургия (второй по мощности в Италии металлургич. комбинат Корнильяно), нефтепереоаботка (1/10 всех мощностей страны). Имеются текст., джутовые, химич., пищевые и др. предприятия. Несколько ТЭЦ. Ун-т (15 в.). Г. - центр туризма. Т. А. Галкина. В городе, живописно расположенном амфитеатром по склонам вокруг бухты, много памятников архитектуры, древнейшие из к-рых - церковь Санта-Мария ди Кастелло (11 в.) и собор Сан-Лоренцо (начат в 11 в., освящён в 1118, позднее перестраивался). Гл. достопримечательность Г. - дворцы и виллы 16-17 вв. (Палаццо Пароли, 1567, виллы Камбьязо, 1548, и Паллавичино делле Пескьере, 1560-72, - арх. Г. Алесси, Палаццо Муничипале, или Дориа-Турси, 1564, арх. Р. Лураго; Палаццо Дураццо-Паллавичини, 1618, и Палаццо дель Университа, 1634-36, - арх. Б. Бьянко) с фонтанами, арочными галереями дворов, лестницами и террасными парками, в композиции к-рых эффектно использован круто падающий рельеф. Для Г. характерен резкий контраст пышной застройки гл. улиц и соседствующих с ними трущобных р-нов. В тесноту старых кварталов вписываются новые здания и комплексы (территория Международной выставки с овальным спортивно-выставочным залом диам. 112 м, 1964, арх. К. Данери и др.; высотное здание Итал. телефонной компании, 1969, арх. М. Бега, П. Гамбачано). Р-ны массового жил. строительства находятся гл. обр. за чертой города (р-н Бернабо-Бреа, 1951-54, арх. К. Данери и др.); дорогие участки на берегу моря застраиваются частными и кооперативными домами-люкс с обширными, роскошно отделанными квартирами (напр., дом на пляже, 1952, арх. К. Данери). Музеи: Гал. Палаццо Россо, Гал. Палаццо Бьянко, Гал. Палаццо Спинола, Гор. музей лигурийской археологии (осн. в 1892), Музей лигурийской архитектуры и скульптуры, подземный музей-сокровищница собора Сан-Лоренцо (1956, арх. Ф. Альбини). А. И. Опочинская. Историческая справка. В древности - поселение лигуров. Завоёванная в 3 в. до н. э. римлянами, была одним из важнейших торг, портов Рим. гос-ва. В период раннего средневековья экономич. значение Г. упало. При лангобардах (с 641) Г. - центр герцогства, а с 9 в. - маркграфства. С 10 в. в городе укрепляется власть епископа. В 11 в., после освобождения от арабов сев. части Тирренского моря, Г. завязала обширные торг, связи с Юж. Италией, Сицилией, Испанией, Африкой. Участие Г. в крестовых походах способствовало превращению её в могущественную мор. державу. В 12 в. Г. - коммуна, власть в к-рой сосредоточилась в руках крупных купцов и зем. магнатов, участвовавших в торговле. Во главе коммуны сначала стояла коллегия консулов, с 1217 - подеста, с 1257 - капитан народа, с 1270 - 2 капитана народа, с 1339 - дож. Гл. соперниками Г. были мор. республики Пиза и Венеция. В 1284 Г. разгромила флот Пизы при Мелории, а в 1298 - флот Венеции при Курцоле, что усилило позиции Г. в Византии и в вост. части Средиземного моря, а также в Чёрном море; в 13-14 вв. Г. владела мн. колониями в Крыму (Кафа и др.). В 1380 Г. потерпела от Венеции поражение при Кьодже. Силы Г. были сломлены; с 1396 она неоднократно попадала под власть Франции и Милана. В 1528 была восстановлена республика Г., зависимая от Испании. В Г. утвердилась олигархия знати. Потеря Черномор, колоний в 15 в., перемещение торг. путей в Атлантич. ок. в 16 в. привели к сокращению торг, значения Г. и свёртыванию судостроения, в к-ром ещё в 14 в. появились зачатки капиталистич. отношений. В 15-16 вв. в Г. процветала шёлкодельческая пром-сть, однако гл. видом хоз. активности Г. стало банковское дело и прежде всего кредит, операции. Гл. финанс. центром Г. стал банк Сан-Джорджо, занимавшийся междунар. кредит, операциями. В 1797-1805 терр. Генуэзской республики составляла Лигурийскую республику. В Г., входившей с 1815 в Сардинское королевство, в 1849 вспыхнуло респ. восстание, подавленное королев, войсками. Со 2-й пол. 19 в. Г., превратившаяся в крупный пром. город, стала важным центром рабочего движения. В 1892 в Г. была осн. Итал. социалистич. партия. Во время 2-й мировой войны Г., оккупированная в сент. 1943 нем.-фаш. войсками, являлась одним из значит, опорных пунктов Движения Сопротивления. Была освобождена партизанами и восставшим населением в апр. 1945. После войны в муниципальных органах Г. сильны позиции левых сил (до 1951 во главе муниципальных органов Г. стояли коммунисты). Генуя. Башня ворот Порта Сопрана. 1155. Генуя. Собор Сан-Лоренцо. 11 - 16 вв. Лит.: Vitаlе V., Breviario della storia di Genova, v. 1 - 2, Genova, 1955; Rutenburg V. I., Gli Uzzano e Genova, в кн.: Miscellaneastorica ligure, fasc. 3, Milano,1963; Соzzani E., Genova, 2 ed., Torino, 1961. В. И. Рутенбург. ГЕНЦ (Gentz) Фридрих (2. 5. 1764, Бреслау, - 9. 6. 1832, Вейнхаус, около Вены), австрийский политич. Деятель и публицист. Род. в семье прусского чиновника. С 1786 на прусской гос. службе, в 1802 перешёл на австрийскую. С сер. 90-х гг. 18 в. в качестве публициста яростно выступал против франц. революции, затем против наполеоновской Франции. Получал субсидии от ряда стран (в т. ч. с 1802 от Англии). Близкий советник и доверенное лицо К. Меттерниха, Г. был секретарём Венского конгресса 1814-15, конференции союзных министров в Париже в 1815, конгрессов Священного союза в Ахене, Вероне, Лайбахе и Троппау. Был одним из активных защитников феод.-монархии, реакции. Сочинения Г. - важный ист. источник. Соч.: Ausgewählte Schriften..., Bd 1 - 5, Lpz., 1836 - 38; Tagebücher..., Bd 1 - 4, Lpz., 1873-74; Tagebücher (1829-1831), W.,[1921]; Briefe, Bd 1 - 3, Munch. - В., 1909 - 13. Лит.: Sweet P. R., Friedrish von Gentz. Defender of the Old Order, Madison, [1941]; Mann G., Friedrich von Centz. Geschichte eines europäischen Staatsmannes, Z. - W., 1947. А. Б. Герман. ГЕНЦИ ЗАГОВОР, организован в Берне в 1749 С. Генци (S. Henzi), Д. и Г. Фуэтерами среди владельцев мануфактур, торговцев, ремесленников, студентов (всего ок. 70 чел.). Цель Г. з. - свержение гор. патрициата и передача власти бюргерским семьям, введение законодательства о цехах и др. План заговорщиков летом 1749 был выдан властям. 17 июля 1749 Генци и 2 др. организатора заговора были казнены. Ю. П. Мадор. ГЕНЦИАНА (Gentiana), название декоративных видов растений рода горечавка, употребляемое в цветоводстве. ГЕНЦИАНВИОЛЕТ, лекарственный препарат, противоглистное и антисептическое средство. Применяют внутрь при глистных заболеваниях - энтеробиозе и стронгилоидозе, наружно - в растворах или мазях при пиодермии, сикозе и нек-рых др. заболеваниях кожи. Г. широко применяется для окраски препаратов в гистологич. и гл. обр. микробиологич. практике. ГЕО... (от греч. ge - Земля), часть сложных слов, указывающая на их отношение к наукам о Земле, земному шару в целом, земной коре (напр., география, геология). ГЕОАКУСТИКА (от гео... и акустика), раздел акустики, в к-ром изучаются звуковые, инфразвуковые и ультразвуковые явления, происходящие в земной коре. Сюда относятся как природные процессы (напр., акустич. предвестники землетрясений), так и явления, связанные с применением упругих волн для изучения строения и свойств верхних слоев земной коры (акустич. разведка, сейсмич. разведка, глубинное сейсмич. зондирование, ультразвуковая эхолокация). Акустич. разведка производится на определённой, заданной частоте и методом отражения и прозвучивания обнаруживает инородные рудные тела в массивах между водонаполненными скважинами. Акустич. разведка возникла почти одновременно с гидролокацией и эхолокацией морского дна. Эти методы явились первым применением ультразвука для практич. целей. Однако большое поглощение высоких частот (20 кгц) в земной коре ограничивает глубину прозвучивания пород неск. десятками м. При низких звуковых и инфразвуковых частотах глубина прозвучивания повышается, но уменьшается возможность более детального изучения разреза. Большей глубины прозвучивания (до неск. км) удалось достигнуть в результате применения методов сейсмической разведки. Изучение строения слоистой среды для более детального расчленения разреза производится также в самих скважинах (звуковой каротаж). ГЕОАНТИКЛИНАЛЬ (от гео... и антиклиналь) (геол.), частное поднятие земной коры в пределах геосинклинальной системы. Г. представляют собой полосы шириной до неск. десятков и длиной до сотен км. Г. существуют нередко в течение неск. геологич. периодов. На завершающих стадиях развития геосинклинали становятся ядрами складчатых горных сооружений. Примером современной Г. может быть островная дуга Курильских о-вов, древней - хр. Уралтау в осевой части Урала. ГЕОБОТАНИКА (от гео... и ботаника), наука о растительном покрове Земли как совокупности растительных сообществ (фитоценозов). Термин Г. предложил нем. географ растений А. Гризебах (1866) для обозначения географии растений. Полного единства в понимании Г. до сих пор нет. Среди отечественных ботаников одни понимают её как синоним фитоценологии (В. В. Алёхин, В. Н. Сукачёв, А. П. Шенников), др. (В. Б. Сочава) включают в это понятие также всю ботаническую географию. Ещё более широкое понимание Г. с включением в неё экологии растений распространено среди учёных нек-рых зарубежных стран. В начальный период развития Г. осн. внимание уделяли видовому составу фитоценозов, их обусловленности внешней средой и развитию теоретич. представлений о характерных особенностях фитоценоза. Польский ботаник И. К. Пачоский (1891) называл эту науку флорологией, а позднее-фитосоциоло-гией (назв. употребляется преим. в зарубежной лит-ре). С оформлением Г. как самостоятельной науки во 2-й пол. 19 - нач. 20 вв. в ней определились два раздела: общая и специальная Г. Общая Г. изучает гл. обр. закономерности строения фитоценозов, выражающиеся в видовом составе, количеств, отношениях между видами в вертикальном (ярусность) и горизонтальном (мозаичность) расчленении, в наличии экологически сходно специализированных и относительно обособленных групп растений (синузий), во взаиморасположении особей различных видов, наконец, в возрастном составе видовых популяций. Изучение двусторонней зависимости между строением фитоценозов и средой составляет также одну из задач общей Г. Др. задачей общей Г. является изучение периодич. (в т. ч. сезонных) и практически необратимых (т. н. сукцессии) изменений фитоценозов во времени. Разработка принципов классификации фитоценозов составляет ещё одну из важнейших задач общей Г. Глубина, содержательность и общность закономерностей, устанавливаемых общей Г., в большой мере определяются успехами специальной Г. Задача последней состоит в изучении конкретных участков растит, покрова, выявлении разнообразия фитоценозов на них, фактической картины их географич. размещения. В связи с этим разрабатываются первичные (местные) классификации фитоценозов, намечаются осн. направления их зависимости от внешних условий и тенденции изменений фитоценозов во времени. Соответственно специфике объектов и необходимости в связи с этим применения особых методов исследования в специальной Г. б. или м. обособились такие дисциплины, как лесоведение, луговедение, болотоведение и др. В истории Г. можно выделить 3 этапа. В течение первого - от кон. 18 в. до кон. 19 в.- вырабатывалось понятие о фитоценозах как особых природных объектах и накапливались первоначальные сведения об их строении, связях со средой и разнообразии. К самому началу 19 в. относятся высказывания нем. натуралиста А. Гумбольдта о растительном покрове как особом элементе природы. Несколько позже швейц. ботаник О. Декандоль высказал важные для Г. положения о борьбе за существование у растений и о влиянии одних растений на др. Эти представления в более развитой форме впоследствии вошли в определение понятия фитоценоза. Наконец, существенной основой Г. стали результаты изучения лугов и лесов, имевшие практич. направленность. На втором этапе (конец 19 в. - начало 20 в.) ведущее место в Г. принадлежало разработке методов описания фитоценозов и основ их классификации. К этому периоду относятся: определение 3-м (Брюссельским) ботанич. конгрессом (1910) ассоциации как основной классификационной единицы растит, покрова, уточнение определения признаков фитоценозов, разработка методов их изучения, попытки использовать данные измерений, наконец, первые опыты применения к изучению фитоценозов статистич. метода. В это время в Г. интенсивно разрабатывается учение о взаимоотношениях фитоценозов и среды, о причинах и направленности сукцессии, о коренных изменениях растит, покрова в связи с изменениями климата, в частности в связи с оледенениями в Сев. полушарии. Для этого этапа развития Г. характерны возникновение и развитие неск. геоботанич. школ, существующих и ныне: русской, франц.-швейцарской, англо-американской и скандинавской. Каждая из них отличается преим. разработкой тех или иных проблем Г., своеобразной трактовкой осн. единиц растительности, особым подходом к классификации фитоценозов. Третий этап развития Г. начинается с 30-х гг. 20 в., когда критерием отличия фитоценоза от неск. совместно произрастающих растений признано взаимодействие растений, составляющих фитоценоз. Была разработана первая классификация форм влияния растений друг на друга (В. Н. Сукачёв, 1956). Наряду с традиционной борьбой за свет изучаются различные формы корневой конкуренции, аллелопатия и т. д. Благодаря трудам сов. геоботаника Т. А. Работнова население каждого вида в составе фитоценоза всё чаще рассматривают как популяцию, состояние и перспективы существования к-рой в данном ценозе в значит, мере определяются её возрастным составом. Растит, ценоз рассматривается как сложная система, выполняющая функцию планетарного масштаба. Эта функция состоит в более полной, чем доступно одновидовой популяции, аккумуляции солнечной энергии и в обеспечении многократного использования элементов минерального питания в круговороте веществ на Земле. В связи с этим оказались необходимыми изучение взаимозависимости растений друг с другом, а также с животными и микроорганизмами, населяющими фитоценоз, и исследования взаимодействий биоценоза со средой его жизни. На совр. этапе развития Г. широкое распространение, особенно за рубежом, получило учение о растит, покрове как о непрерывном целом - континууме. Для совр. Г. характерно развитие геоботакического картографирования обширных территорий. Основополагающую роль в этом сыграли работы, начатые в 20-х гг. 20 в. под рук. Н. И. Кузнецова. Геоботанич. исследования организуются в зависимости от их конкретной задачи: то как маршрутно-полевые, то как стационарные (в естеств. ценозах и в культурных посевах и посадках). Для описания фитоценозов широко применяется метод пробных площадей (участков) такого размера, чтобы каждая из них отражала осн. свойства фитоценоза в целом. Наряду с этим прибегают к описанию мелких площадок, совокупность к-рых должна статистически достоверно охарактеризовать ценоз. Разрабатываются и способы точного количеств, учёта, напр., надземной и подземной массы, относительной площади светопользования растений и пр. Для более глубокого проникновения в жизнь фитоценоза прибегают к изучению составляющих его компонентов средствами физиологич. исследований, применимых в полевой обстановке, а также к фитоценологич. экспериментам. Как и в др. разделах ботаники, в Г. важное значение имеет сравнительный метод, применяемый прежде всего для объединения фитоценозов в классификационные единицы разных категорий, что необходимо для обозримости материала, для оценки (в т. ч. хозяйственной) территории, её геоботанич. районирования и картирования. Сравнительный метод необходим также и при исследованиях, проводимых в рамках концепции непрерывности (континуальности) растит, покрова. Широкое применение статистич. методов приблизило Г. к математич. моделированию, к-рое ещё не получило широкого распространения в Г., но должно сыграть существенную роль в разработке способов управления фитоценозами с кибернетич. позиций. Г. тесно связана с рядом наук о Земле - с физической географией, метеорологией, гидрологией, климатологией, почвоведением, поскольку фитоценозы в своём составе и строении существенно зависят от внешней среды и сами оказывают на неё глубокое воздействие. Ещё более тесна связь Г. с циклом ботанич. дисциплин, особенно с морфологией (жизненные формы), систематикой, экологией и географией растений. Вопросы истории растит, покрова сближают Г. с историч. геологией, историч. географией, филогенией растений и с палеоботаникой. Г. тесно связана также с рядом агрономич. дисциплин, в частности с луговодством, лесоводством и пр. Г. широко применяется в хоз-ве мн. стран. Учёт, определение площади, установление продуктивности природного растит, покрова и возможностей его улучшения имеют важное практич. значение и связаны с выделом и организацией территорий совхозов и колхозов, с освоением малообжитых р-нов, в частности в тундровой и пустынной зонах. Большое участие принимали геоботаники в разработке теории и в практич. осуществлении проектов полезащитных лесонасаждений. В поле зрения Г. всё шире включаются агрофитоценозы полей, сеяных и полукультурных лугов. В СССР геоботаники входят во Всесоюзное ботаническое общество; участвуют в международных ботанических конгрессах. Достижения Г. в СССР отражаются в Ботаническом журнале (с 1870), в Бюллетене Московского общества испытателей природы. Отдел биологический (с 1887), в Геоботанике (АН СССР. Ботанический институт им. В. Л. Комарова. Труды. Серия 3) (с 1932) и аналогичных изданиях союзных республик и в трудах науч.-исследовательских учреждений, в Лесоведении (с 1967). Важное международное значение имеют зарубежные журналы как общеботанические, так и специализированные [напр., Journal of Ecology (L.- Camb., с 1913), Ecology (N. Y., с 1920), Pflanzensoziologie (Jena, с 1931), Vegetatio (The Hague, с 1949, орган междунар. геоботан. ассоциации), Excepta botanica (Stuttg., с 1959, журнал по ботанич. картографии)]. Лит.: Пачоский И. К., Основы фитосоциологии, Херсон, 1921; Сукачёв В. Н., Растительные сообщества, 4 изд., Л.- М., 1928; Раменский Л. Г., Введение в комплексное почвенно-геоботаническое исследование земель, М., 1938; Морозов Г. Ф., Учение о лесе, 7 изд., М.- Л., 1949; Быков Б. А., Геоботаника, 2 изд., А.-А., 1957; Полевая геоботаника, т. 1 - 3, М.- Л., 1959-64; Ярошенко П. Д., Геоботаника, М.- Л., 1961; Основы лесной биогеоценологии, М., 1964; Шенников А. П., Введение в геоботанику, Л., 1964; Василевич В. И., Статистические методы в геоботанике, Л., 1969; Ярошенко П. Д., Геоботаника. [Учебное пособие], Л., 1969 (библ. с. 195-98); Gams H., Prinzipienfragen der Vegetationsforschung, Z., 1918; Du Riеtz G. E., Vegetationsforschung auf soziationsanalytischer Grundlage, в кн.: Handbuch der biologischen Arbeitsmethoden, Abt. 11, Bd 5, H. 2, В.- W., 1930; Ludi W., Die Methoden der Sukzessionsforschung in der Pflanzensoziologie, в кн.: Handbuch der biologischen Arbeitsmethoden, Abt. 11, Bd 5, H. 3, В.-W., 1930; Rübel E.. Pflanzengesellschaften der Erde, Bern, 1930: Weaver J. E. and С1ements F. E., Plant ecology, 2 ed., N. Y.- L., 1938; Кnapp R., Experimented Soziologie der höheren Pflanzen, Bd 1, Stuttg., 1954; Кlika J., Nauka о rostlinnych spolecenstvech (Fytocenologie), Praha, 1955; Scamoni A., Einführung in die praktische Vegetationskunde, 2 Aufl., Jena, 1963; Вгaun-Blanquet J.. Pflanzensoziologie, 3 Aufl., W.- N. Y., 1964. А. А. Уранов. ГЕОБОТАНИЧЕСКИЕ КАРТЫ, карты растительности, карты, отображающие географич. распространение типологич. подразделений растительности (ассоциаций, групп ассоциаций, формаций), а также их пространственных комбинаций (комплексов, сочетаний, рядов). В зависимости от целевого назначения и принципов построения Г. к. делят на универсальные и специализированные. Универсальные Г. к. показывают распределение естественных единиц растит, покрова, сложившихся в процессе его историч. развития,- коренных растит, сообществ, напр, еловых лесов, ковыльных степей и т. д. На универсальных картах отражаются также все те изменения, к-рым подверглась растительность под влиянием деятельности человека, - кратковременно- и длительно-производные сообщества, напр, берёзовые леса на месте ельников, с.-х. земли на месте ковыльных степей. Специализированные Г. к. отображают черты растительности, наиболее существенные для того или иного направления хоз. её использования, имеют различные прикладные задачи (карты кормовые, лесные, индикационные, растит, ресурсов) и содержат дополнит, показатели, в т. ч. и количественные. Универсальные Г. к. в зависимости от масштаба подразделяются на детальные крупномасштабные (1 : 5000 - 1 : 25 000), обобщённые крупномасштабные (1 : 50 000-1 : 200 000), среднемасштабные ( 1: 300 000-1 : 1 000 000), мелкомасштабные формационные (1:1 500 000- 1 : 4 000 000), мелкомасштабные обзорные (1 : 5 000 000 и мельче). Новейшие обзорные мелкомасштабные Г. к. мира, частей света и СССР помещены в Физико-географическом атласе мира (1964); наиболее подробная из опубликованных Г. к. СССР имеет масштаб 1 : 4 000 000 (1954). Г. к. отдельных стран и их частей входят во многие комплексные атласы, например Atlas de France (1954), атласы Узб. ССР (1963), Азерб. ССР (1963), Забайкалья (1967), а также выпускаются отд. изданиями. Специализированные Г. к. (кормовые, лесные, сырьевых ресурсов) помещены во мн. атласах - Ленинградской (1967), Кустанайской (1963) областей и др. (Образец геоботанической карты см. на вклейке к стр. 256.) Лит.: Принципы и методы геоботаничеcкого картографирования, М.- Л., 1962; Геоботаническое картографирование, М.- Л., 1963 - 68; Kuchler A. W., Vegetation mapping, N. Y., 1967. Т. И. Исаченко. ГЕОГЕЛЬМИНТЫ (от гео... и гельминты), группа паразитич. червей человека и животных, развивающихся (в отличие от биогельминтов) без промежуточных хозяев. Яйца Г. с фекалиями попадают в почву, где развиваются в тёплое время года до стадии личинок. Заражение человека происходит либо через немытые овощи, фрукты, руки, на к-рых находятся инвазионные яйца (напр., аскариды, власоглава человеческого, острицы), либо при непосредственном контакте с землёй, где живут личинки (напр., анкилостомид). ГЕОГРАФА ЗАЛИВ (Geographe Bay), залив Индийского ок. у юго-зап. берега Австралии. Вдаётся в сушу на 65 км. Глуб. до 27 м. Берега окружены песчаными отмелями и рифами. Впадает р. Васса. Приливы суточные, их величина 1 м. На В., у входа в залив, - порт Банбери, на Ю. - город и порт Басселтон. Г. з. открыт в 1801 франц. экспедицией под нач. капитана Н. Бодена на кораблях Географ и Натуралист. Назван в честь корабля. ГЕОГРАФИИ ИНСТИТУТ Академии наук СССР (ИГАН), научно-исследоват. ин-т, разрабатывающий теоретич. и практич. вопросы в области физич. и экономич. географии. Возник в 1918 в виде Промышленно-геогр. отдела в составе Комиссии по изучению естеств. производит, сил (КЕПС) АН СССР в Петрограде. В 1930 отдел был преобразован в Геомопфологич. ин-т АН СССР, к-рый в 1934 был переведён в Москву и затем переименован в Институт физич. географии, а в 1936 - в Институт географии. Организатором и руководителем (до 1951) института был акад. А. А. Григорьев. ИГАН имеет (1971) отделы: физич. географии и геофизики ландшафтов, геоморфологии и палеогеографии, климатологии, гидрологии, гляциологии, внутр. водоёмов, биогеографии, географии почв и геохимии ландшафтов, экономии, географии СССР и др. социалистич. стран, географии капиталистич. и развивающихся стран, территориально-экономич. проблем, картографии, а также Курскую экспериментальную полевую станцию, спорово-пыльцевую, стереофотограмметрическую и др. лаборатории, Главным направлением деятельности ИГАН служит разработка взаимосвязанных проблем преобразования природы, направленных на эффективное использование естеств. ресурсов, и научного прогнозирования ожидаемых изменений природных условий, а также экономико-геогр. проблем развития территориально-производств. комплексов, экономич. оценки крупных проектов преобразоват. мероприятий. С этой тематикой, имеющей научно-практич. направленность, органически связана разработка общих теоретич. вопросов, существенно влияющих на развитие системы геогр. наук. ИГАН организует экспедиции в различные р-ны СССР, участвует в междунар. конгрессах и совещаниях. С 1931 по I960 издавал Труды Ин-та географии АН СССР. Публикует монографии по физ. и эконом, географии СССР и зарубежных стран, а также серию Природные условия и естественные ресурсы СССР. Лит.: Каманин Л. Г., Рихтeр Г. Д., Фрадкин Н. Г., Из полувековой истории Института географии АН СССР, Изв. Академии наук СССР. Серия географическая, 1968, № 6; Издания Института географии АН СССР. Библиографический указатель, М., 1959; Известия АН СССР. Серия географическая. Систематический указатель (1951 - 1966), М., 1967. М. И. Нейштадт. ГЕОГРАФИИ СИБИРИ И ДАЛЬНЕГО ВОСТОКА ИНСТИТУТ Сибирского отделения АН СССР (ИГС и ДВ), организован в 1959 в г. Иркутске. Занимается изучением комплексных географич. проблем, а также вопросов региональной и прикладной географии (освоение таёжных территорий, принципы районирования и классификации земель на ландшафтной основе и пр.). Исследуются вопросы формирования населения, медицинской географии, географии производства и оценки природных ресурсов. К задачам ин-та относятся разработка принципов и методов тематич. картографирования (ландшафтного, медико-географического, геоботанического и др.) и составление тематических карт. На стационарах ин-та ведутся экспериментальные физико-географич. наблюдения и исследуются природные режимы таёжных и степных ландшафтов для целей прогнозирования изменений природной среды и выявления теоретич. вопросов ландшафтоведения. В составе ин-та имеются лаборатории и геогр. стационары в Зап. и Вост. Сибири. В Чите - с 1963 региональная лаборатория ИГС и ДВ; в пос. Шушенское (Красноярский край) с 1968 - Южносибирская географическая обсерватория ин-та. С 1962 публикуются Доклады ин-та географии Сибири и Дальнего Востока и ежегодник Сибирский географический сбооник, а также различные издания по физич. и экономич. географии. Лит.: Воробьева Т. Н., Институту географии Сибири и Дальнего Востока - 10 лет, Изв. Всесоюзного географического об-ва, 1970, т. 102, в. 3. В. Б. Сочава. ГЕОГРАФИЧЕСКАЯ ДОЛГОТА, см. Географические координаты. ГЕОГРАФИЧЕСКАЯ ОБОЛОЧКА, ландшафтная оболочка, эпигеосфера, оболочка Земли, в которой соприкасаются и взаимодействуют литосфера, гидросфера, атмосфера и биосфера. Характеризуется сложным составом и строением. Верхнюю границу Г. о. целесообразно проводить по стратопаузе, т. к. до этого рубежа сказывается тепловое воздействие земной поверхности на атмосферные процессы; границу Г. о. в литосфере часто совмещают с нижним пределом области гипергенеза (иногда за нижнюю границу Г. о. принимают подножие стратисферы, среднюю глубину сейсмических или вулканич. очагов, подошву земной коры, уровень нулевых годовых амплитуд температуры). Таким образом, Г. о. полностью охватывает гидросферу, опускаясь в океане на 10-11 км ниже поверхности Земли, верхнюю зону земной коры и нижнюю часть атмосферы (слой мощностью 25-30 км). Наибольшая толщина Г. о. близка к 40 км. Качественные отличия Г. о. от других оболочек Земли: Г. о. формируется под действием как земных, так и космических процессов; исключит, богата разными видами свободной энергии; вещество присутствует во всех агрегатных состояниях; чрезвычайно разнообразна степень агрегированности вещества - от свободных элементарных частиц через атомы, ионы, молекулы до химич. соединений и сложнейших биологич. тел; концентрация тепла, притекающего от Солнца; наличие человеческого общества. Основные вещественные компоненты Г. о. - это слагающие земную кору горные породы (с их формой - рельефом), воздушные массы, водные скопления, почвенный покров и биоценозы; в полярных широтах и высокогорьях существенна роль скоплений льда. Основные энергетич. компоненты - гравитационная энергия, внутреннее тепло планеты, лучистая энергия Солнца и энергия космич. лучей. При всей ограниченности набора компонентов сочетания их могут быть весьма многообразными; это зависит и от числа входящих в сочетание слагаемых и от их внутренних вариаций (поскольку каждый компонент - это тоже очень сложная природная совокупность), а главное - от характера их взаимодействия и взаимосвязей, т. е. от географич. структуры. Г. о. присущи следующие важные черты: 1) целостность Г. о., обусловленная непрерывным обменом вещества и энергии между её составными частями, поскольку взаимодействие всех компонентов связывает их в единую материальную систему, в к-рой изменение даже одного звена влечёт сопряжённое изменение и всех остальных. 2) Наличие круговорота веществ (и связанной с ним энергии), обеспечивающего многократность одних и тех же процессов и явлений и их высокую суммарную эффективность при ограниченном объёме исходного вещества, участвующего в этих процессах. Сложность круговоротов различна: одни из них - механич. движения (циркуляция атмосферы, система морских поверхностных течений), другие сопровождаются сменой агрегатного состояния вещества (влагооборот на 3емле), в третьих происходит также и его химическая трансформация (биологич. круговорот). Круговороты, однако, не замкнуты, и различия между их начальными и конечными стадиями свидетельствуют о развитии системы. 3) Ритмика, т. е. повторяемость во времени различных процессов и явлений. Она обусловлена гл. обр. астрономич. и геологич. причинами. Выделяется ритмика суточная (смена дня и ночи), годовая (смена времён года), внутривековая (например, циклы в 25-50 лет, наблюдаемые в колебаниях климата, ледников, уровней озёр, водоносности рек н т. п.), сверхвековая (напр., смена за каждые 1800-1900 лет фазы прохладно-влажного климата фазой сухого и тёплого), геологическая (циклы каледонский, герцинский, альпийский по 200-240 млн. лет каждый) и т. п. Ритмы, как и круговороты, не замкнуты: то состояние, какое было в начале ритма, в конце его не повторяется. 4) Непрерывность развития Г. о., как нек-рой целостной системы под влиянием противоречивого взаимодействия экзогенных н эндогенных сил. Следствиями н особенностями этого развития являются: а) территориальная дифференциация поверхности суши, океана и морского дна на участки, различающиеся по внутренним особенностям и внешнему облику (ландшафты, геокомплексы): определяется пространственными изменениями географической структуры: особые формы теориториалыюй дифференциации - географическая зональность (см. Зоны физико-географические) и высотная поясность; б) полярная асимметрия, т. е. существенные оазличия природы Г. о. в Северном н Южном полушариях; проявляется в распределении суши и моря (подавляющая часть суши в Сев. полушарии), климата, состава животного и растительного мира, в характере ландшафтных зон и т. п.; в) гетерохронность (по К. К. Маркову, метахронность) развития Г. о., обусловленная пространственной разнородностью природы Земли, вследствие чего в один и тот же момент разные территории либо находятся в различных фазах одинаково направленного эволюционного процесса, либо отличаются друг от друга направлением развития (примеры: древнее оледенение в разных районах Земли начиналось и кончалось неодновременно; в одних геогр. зонах климат становится суше, в других в то же время - влажнее и т. п.). Г. о. является предметом изучения физической географии. К идее Г. о. впервые подошли П. И. Броунов (1910) и Р. И. Аболин (1914). Самый термин Г. о. ввёл и обосновал А. А. Григорьев (1932). Понятия, аналогичные понятию Г. о., есть и в зарубежной геогр. литературе (земная оболочка А. Гетнера и Р. Хартшорна, геосфера Г. Кароля и др.). Однако там Г. о. рассматривается обычно не как природная система, а как совокупность природных и обществ, явлений. Лит.: Аболин Р. И.. Опыт эпигенологической классификации болот, Болотоведение, 1914. № 3; Броунов П. И., Курс физической географии, П., 1917; Григорьев А. А., Опыт аналитической характеристики состава и строения физико-географической оболочки земного шара, Л. - М., 1937; его же, Закономерности строения и развития географической среды, М., 1966; Исаченко А. Г., Системы и ритмы зональности, Изв. Всесоюзного географического об-ва, 1971, т. 103, в. 1; Калесник С. В., Общие географические закономерности Земли, М., 1970; Марков К. К., Полярная асимметрия географической оболочки, Изв. Всесоюзного географического общества, 1963, т. 95, в. 1; его же, Пространство и время в географии, Природа, 1965, № 5; Carol H., Zur Theorie der Geographie, Mitteilungen der Oesterreichischen Geographischen Gesellschaft, 1963, Bd 105 H. 1-2; Hartshоrne R., The nature of geography, Lancaster, 1939. С. В. Колесник. ГЕОГРАФИЧЕСКАЯ ОСНОВА карты, общегеографич. элементы тематич. карты, не входящие в её специальное содержание. При пользовании картой Г. о. облегчает ориентирование и уяснение закономерностей размещения явлений, относящихся к тематике карты. Тема и назначение карты определяют содержание и характер Г. о.: на мелкомасштабных тематич. картах мира и материков содержание Г. о. часто ограничивается изображением береговой линии, крупнейших рек и озёр, важнейших городов; на картах более крупных масштабов Г. о., помимо этого, может включать растительный покров, рельеф, пути сообщения, политич. и адм. границы, ледники, болота и др. Особенно детальна и наглядна Г. о. на картах, предназначенных для практич. использования, где она обеспечивает пространственную локализацию изображённых явлений. При составлении тематич. карт Г. о. служит своего рода каркасом для построения картографич. изображения, нанесения и укладки спец. содержания. При подготовке серийных тематич. карт и атласов широко применяется изготовление типовой Г. о., к-рая составляется и оформляется один раз и используется для ряда взаимосвязанных карт. Ю. Г. Кельнер. ГЕОГРАФИЧЕСКАЯ ПАТОЛОГИЯ, область медицины, изучающая патологию человека, животных и растений в связи с географич. факторами, а для человека также и с социально-экономическими. Г. п. исследует процессы взаимодействия организма и среды обитания, характер клинич. проявления болезней в связи с местными особенностями природы, образа жизни населения и социальными факторами. Г. п. тесно соприкасается с географией населения, антропологией, экологией, демографией, социальной гигиеной, гигиеной коммунальной, гигиеной питания и гигиеной труда. Термин Г. п. был предложен в 1858 нем. патологом, эпидемиологом и гигиенистом А. Хиршем. В 1929-31 нем. учёным М. Асканази было основано Международное общество Г. п., ставившее своей целью сопоставление патологич. панорамы в различных странах мира. К 1969 состоялось 9 конференций этого общества, материалы к-рых печатаются в швейц. Журнале патологии и микробиологии. В СССР Г. п. зародилась и развивалась под назв. краевой патологии. Сов. учёным Е. Н. Павловским было создано учение о природной очаговости инфекционных болезней, а Л. А. Зильбером, М. П. Чумаковым, А. К. Шубладзе, Е. Н. Левкович, В. Д. Селовьёвым, А. А. Смородинцевым, А. В. Чуриловым, М. К. Кронтовской, Н. Н. Сиротининым и др. были открыты такие природноочаговые болезни человека, как клещевой энцефалит, геморрагич. нефрозонефрит и группа геморрагич. лихорадок, клещевой сыпной тиф Сев. Азии, пароксизмальный риккетсиоз, алиментарно-токсическая алейкия, гелиотропный гепатит с асцитом, отравление ядовитым сорняком (седой триходесмой) в Ср. Азии, алиментарная миоглобинурия, связанная с употреблением в пищу нек-рых видов рыб, молибденовая подагра, стронциевая хондро-дистрофия и др. Сов. учёные определили районы СССР, где были обнаружены инфекционные, паразитарные или биогеохимические заболевания, первоначально открытые за рубежом. Г. п. в СССР отличается от Г. п. капиталистич. стран органич. связью с организацией здравоохранения, социальной гигиеной и географией медицинской. Г. п. изучает все проявления нарушенной или изменённой жизнедеятельности организма, определяемые лишь спец. исследованиями (клиническими, биохимическими, патофизиологическими или патоморфологическими), в то время как нозогеография (география болезней) учитывает только выраженные болезни. Таким образом, Г. п. регистрирует и исследует не только развившиеся болезни или их самые начальные стадии, но и предболезни, т. е. те нарушения в организме, к-рые рано или поздно приводят к заболеванию. Для территории СССР, характеризующейся исключительным разнообразием условий внешней среды, Г. п. имеет особенно важное значение, тем более, что условия жизни в обширных регионах (напр., Арктика, пустыни и полупустыни, высокогорья, территории, отличающиеся повышенной сейсмичностью, и т. п.) трудны для приспособляемости человеческого организма. В этих регионах особенно часто встречаются своеобразные реакции организма на среду обитания и необычные формы клинич. течения болезней. Г. п. подробно исследует данные по злокачественным опухолям, сердечно-сосудистым и наследственным болезням, к-рые неравномерно встречаются на территориях разных стран. Это объясняется не только наличием исторически сложившихся изолятов или обычаями населения, но и тем, что нек-рые профессиональные вредности (радиоактивные и химич.) могут вызывать стойкие наследственные изменения в половых и соматич. клетках - мутации. Сов. учёные исследуют Г. п. и др. стран, что имеет важное значение при тесных международных связях вследствие возможности заноса возбудителей или переносчиков уже ликвидированных или вообще никогда не встречавшихся в СССР болезней. Природная очаговость болезней существует также у животных и растений. Эти болезни могут быть не только инфекционного, но и биогеохимического происхождения. Напр., дефицит или избыток нек-рых микроэлементов в почве вызывает заболевания растений и животных - т. н. биогеохимические эндемии (см. Биогеохимия). Вопросы Г. п. периодически освещаются в реферативном журнале , Медицинская география, выпускаемом Всесоюзным институтом научной и технической информации (ВИНИТИ) АН СССР. Организацией исследований по Г. п. за рубежом занимается Международное общество географической патологии. Лит.: Авцын А. П., Предмет, задачи и методы советской географической патологии, Вестник АМН СССР, 1964, № 12; Hirsch A., Handbuch der historisch-geographischen Pathologie, Abt. 1-3, Stuttg., 1881-86; Henschen F., Grundzüge einer historischen und geographischen Pathologie, В.- [u.a.], 1966. А. П. Авцын. ГЕОГРАФИЧЕСКАЯ СРЕДА, часть земного природного, в той или иной степени изменённого людьми окружения человеческого общества, с которой общество в данный момент непосредственно связано в своей жизни и производственной деятельности. У Г. с. четыре основных признака: 1) Г. с.- земное окружение общества; даже если человечество выйдет за пределы Земли, оно не сможет унести с собой Г. с.; на других планетах оно встретится с другой средой, а не с географической. 2) Г. с.- природное окружение человеческого общества, т. е. комплекс природных условий, возникших независимо от человека и сохранивших, несмотря на воздействие на них людей, способность к дальнейшему саморазвитию по законам, действующим в географической оболочке Земли; следовательно, элементы среды, созданные из природных веществ трудом и сознательной волей человека, но лишённые дальнейшего саморазвития и не имеющие аналогов в девственной природе, в состав Г. с. уже не входят и образуют особую - техногенную среду общества (города, заводы, электростанции и т. п.), сосуществующую и тесно взаимодействующую с Г. с. 3) Г. с.- сфера непосредственного взаимодействия природы и общества; следовательно, территории, лежащие вне этой сферы, к Г. с. не относятся, хотя последствия производственной деятельности человечества (напр., общее повышение содержания СО2 в земной атмосфере, выпадение радиоактивных осадков после атомных взрывов и т. п.) могут и сказываться на их природе косвенным образом. 4) Г. с. расширяется со временем по объёму и содержанию, т. к. для обеспечения круга своих потребностей человеческое общество вовлекает в эксплуатацию всё новые земные пространства, новые стороны и составные части природы; при том же богатстве земной природы Г. с. в прошлом была более ограниченной, чем в настоящее время. Стремление нек-рых учёных отождествить Г. с. с географич. оболочкой Земли ошибочно: Г. с. по мере развития человеческого общества пространственно расширяется, а географич. оболочка - нет; только в будущем Г. с. охватит всю географич. оболочку (совпадёт с ней) и даже выйдет за её границы, не отрываясь, впрочем, от Земли. Географич. оболочка стала Г. с. человеческого общества только в связи с возникновением последнего (ранний палеолит) и только на той территории, на к-рой общество жило и трудилось. В географической лит-ре были попытки отнести к Г. с. и самоё человеческое общество (т. е. сделать его своим собственным окружением), и орудия труда, и все предметы и объекты, созданные руками человека. Эти взгляды, ввиду их необоснованности, у большинства советских географов признанием не пользуются. Г. с.- одно из постоянных и необходимых условий развития общества; она может ускорять или замедлять это развитие, но не является его главной движущей силой, так как специфич. законы движения природы и общества, равно как и темпы этого движения (изменения), существенно различны. Развитие общества определяется способом производства. Эта марксистская концепция раскрыла ошибочность других взглядов на роль Г. с. в развитии общества - географического нигилизма (полное отрицание этой роли), географического детерминизма (Г. с. приписывается определяющее значение), географического поссибилизма (игнорирование характера общественного строя во взаимодействии природы и общества). См. также Географическая школа в социологии. Лит.: Иванов-Омский И. И., Исторический материализм о роли географической среды в развитии общества, М., 1950; Саушкин Ю. Г., Географическая среда человеческого общества, География и хозяйство, 1963, сб. 12; Калесник С. В., Проблема географической среды, Вести. ЛГУ, 1968, в. 12. С. В. Калесник. ГЕОГРАФИЧЕСКАЯ ШИРОТА, см. Географические координаты. ГЕОГРАФИЧЕСКАЯ ШКОЛА в социологии, направление в немарксистской социологии, рассматривающее географическую среду или её отд. компоненты (климат, почву, реки и т. и.) в качестве определяющего фактора развития общества. Идея обусловленности обществ, явлений географич. средой высказывалась в античности (Демокрит, Гиппократ, Геродот, Полибий, Страбон), ср. века (араб, мыслитель Ибн Хальдун), Ж. Боденом во Франции в 16 в. Основателем Г. ш. считают франц. мыслителя Ш. Монтескье, развившего идеи о влиянии географич. условий, климата на жизнь людей, обычаи и нравы народов, на становление хоз. и даже политич. строя различных стран. Видным представителем Г. ш. был англ, историк Г. Т. Бокль. Проблематика Г. ш. исследовалась мн. географами, историками и экономистами конца 19 - 1-й пол. 20 вв. (нем. географ и этнограф Ф. Ратцель, франц. географ Э. Реклю, амер. географ Э. Хантингтон, рус. учёный Л. И. Мечников и др.). Вначале идеи Г. ш., несмотря на их односторонность, были направлены против релит, идеологии и культа великих людей, выдвигали принцип детерминизма в обществ, жизни. Во 2-й пол. 19 в., номере развития научной социологии, Г. ш. утратила своё прогрессивное содержание. Игнорирование социальных движущих сил историч. прогресса и выдвижение географич. среды в качестве решающего фактора в экономич., политич. и культурном развитии народов нередко использовались для обоснования реакц. теорий о природных причинах отсталости колониальных народов, оправдания колониальной политики капиталистич. держав. В эпоху империализма географич. детерминизм у ряда реакц. идеологов привёл к прямой апологии и идейному обоснованию империалистич. экспансии (см. Геополитика). В совр. социологии Г. ш. не существует как самостоят, направление, а поставленные ею проблемы исследуются в соответствующих отраслях социологии (социальная экология и др.). Лит.: Мечников Л. И., Цивилизация и великие исторические реки, пер. с франц., М., 1924; Ковалевский М. М., Социология, т. 1-2, СПБ, 1910. См. также лит. при ст. Геополитика к Географическая среда. В. И. Коровиков. ГЕОГРАФИЧЕСКИЕ ЖУРНАЛЫ, периодич. издания, освещающие вопросы географии. Развитие географической журнальной литературы относится к 18 в. и связано с деятельностью академий наук, геогр. обществ, университетов и специальных научных учреждений. Первый русский Г. ж. Исторические, генеалогические и географические примечания в „Ведомостях" издавался с 1728. В 1865 был основан журнал Изв. Русского географического общества (ныне Известия Всесоюзного географического общества), к-рый выделялся научной ценностью опубликованных в нём материалов, связанных с именами видных русских географов - П. П. Семёнова-Тян-Шанского, А. И. Воейкова, Ю. М. Шокальского и др. С 1894 Московским обществом испытателей природы начал издаваться журнал -Землеведение (с 1940 выходит непериодическими сборниками), основанный Д. Н. Анучиным. В СССР развитие географической науки обусловило появление различных Г. ж. и повременных (непериодических) изданий. Кроме Изв. Всесоюзного географического общества (с 1865), издаются: Изв. АН СССР. Серия географическая (с 1951), Тр. Ин-та географии АН СССР (с 1931), сборники Ин-та географии Сибири и Дальнего Востока Сибирского отд. АН СССР, Изв. АН Азербайджанской ССР. Серия геолого-географических наук (с 1958), Изв. АН Армянской ССР. Серия геологических и географических наук (с 1957), Тр. Ин-та географии им. Вахушти АН Грузинской ССР (с 1947), Вестник Московского университета. Серия 5. География (с 1960), Вестник Ленинградского университета. Геология. География (с 1946), сборники Московского филиала Геогр. об-ва Вопросы географии (с 1946), сборники Вост. комиссии Географич. об-ва Страны и народы Востока (с 1959); филиалы и отделения геогр. общества в др. городах публикуют свои географические сборники. Во многих университетах и педагогич. институтах издаются специальные выпуски Записок и Учёных трудов, посвящённые вопросам географии. Для учителей предназначен журнал География в школе (1934), для молодёжи - популярный Г. ж. Вокруг света. Географич. материалы помещаются также в журн. Геоморфология (с 1970), Океанология (с 1961), Метеорология и гидрология (с 1950), Природа (с 1912), Наука и жизнь (с 1934) и др. За рубежом издаётся ок. 220 периодич. журналов и сборников, в т. ч. во Франции 23, США 22, в Великобритании, ФРГ и Японии по 15. Св. половины (52%) зарубежных географических периодич. изданий приходится на общегеографические научные журналы и сборники, ок. 30% на издания по отд. отраслям географии; менее распространены региональные, научно-педагогич. и научно-популярные издания. Значит, развитие Г. ж. получили в зарубежных социалистич. странах; среди них наиболее известны: в ГДР - Реtermanns geographische Mitteilungen (Gotha, с 1855), Zeitschrift fur den Erdkundeunterricht (В., с 1949); в Болгарии - научно-популярный Г. ж. География (София, с 1950) и Известия на Географския институт Българската Академия на науките (София, с 1951); в Чехословакии - сборники Географического общества Sbprnik Ceskoslovenske Spolecnosti zemepisne (Praha, с 1896), Geograficky casopis (Bratislava, с 1949); в Польше - Przeglad geograficzny (Warsz., с 1918), Czasopismo geograficzne (Lodz - Warsz., с 1923); в Венгрии - Acta geographica. Acta universitatis szegediensis (Szeged, с 1955), Foldrajzi kozlemenyek (Bdpst, с 1873) и др.; в Румынии - Natura (Buc., с 1949); в Югославии - Geografski glasnik (Zagreb, с 1949), Geografski vestnik (Ljubljana, с 1925). В зарубежных социалистич. странах Г. ж. всё больше внимания уделяют разработке научных проблем, диктуемых основными задачами строительства социализма. Из Г. ж., издающихся в капиталистич. странах, наиболее известны: в Великобритании - The Geographical Journal (L., с 1893), Geographical Magazine (L., с 1935); во Франции - Annales de geographic (P., с 1891); в США - Geographical Review (N. Y., с 1916), Annals, of the Association of American Geographers (Wash., с 1911), Professional Geographer (Wash., с 1949), Economic Geography (Worcester, с 1925); в Италии - Rivista geografica italiana (Firenze, с 1893), Annali di ricerche e studi dt geografia (Genova, с 1945); в Швеции - Geografiska annaler (Stockholm, с 1919); в Финляндии-Fennia (Hels., с 1889); в Швейцарии-Geographica helvetica (Z., с 1946); в ФРГ - Berichte zur deutschen Landeskunde (Remagen- Stuttg., с 1941), Erdkunde. Archiv fur Wissenschaftliche Geographic (Bonn, с 1947); в Японии - Тигираку Хёрон (Токио, с 1925) и Тири (Токио, с 1956) и др. Всё большее развитие получают Г. ж. в развивающихся странах; так, в Индии наиболее известны Geographical Review of India (Calc., с 1936),. National Geographical Journal of India (Benares, с 1955); в Бразилии - Boletim geografico (Rio de J., с 1943 - Revista Brasileira de geografia (Rio de J., с 1939); в Мексике - Boletin de la Sociedad mexicana de geografia у estadistica (Мех., с 1839); в ОАР - Bulletin de la Societe de geographic d'Egypte (Le Caire, с 1922); в Нигерии - Nigerian Geographical Journal (Ibadan, с 1957); в Эфиопии - Ethiopian Geographical Journal (Addis Abeba, с 1963) и т. д. В ряде стран выпускаются Г. ж., предназначенные специально для учителей географии [напр., Journal of Geography (Chi.-N. Y., с 1902), Geography (Manchester, 1895-1902)]. В капиталистич. странах издаются также популярные Г. ж., заполняемые гл. обр. фотографиями и пропагандистским материалом, далёким от науки [напр., издаваемый в США National Geographical Magazine (Wash., с 1888)]. И. И. Пархоменко. ГЕОГРАФИЧЕСКИЕ ИНСТИТУТЫ научно-исследовательские, научные учреждения, занимающиеся исследовательской работой .в области географии. Существуют в большинстве крупных стран мира. Наряду с комплексными Г. и. в ряде стран существуют отраслевые Г. и. В СССР крупнейшими являются Географии институт АН СССР (Москва), Географии Сибири и Дальнего Востока институт Сибирского отделения АН СССР (Иркутск); в 1971 создан Тихоокеанский институт географии Дальневост. науч. центра АН СССР. Имеются также ин-ты географии в. АН Азерб. ССР (в Баку), АН Груз. ССР (им. Вахушти, в Тбилиси); в Украинской, Белорусской, Казахской, Литовской, Молдавской, Киргизской и Армянской ССР работают секторы или отделы географии в подразделениях АН этих республик. Геогр. тематикой по соответственным отраслям знаний занимаются также институты водных проблем АН СССР, Азерб. ССР и Арм. ССР, Лаборатория озероведения АН СССР, Ин-т пустынь Туркм. ССР. Значит, геогр. исследования осуществляет Океанологии институт АН СССР. Большую работу проводят геогр. факультеты ун-тов, пед. ин-тов и др. высших учебных заведений, а также Географическое общество Союза ССР. За рубежом ин-ты географии часто существуют при ун-тах и сочетают исследовательскую работу с педагогич. В ГДР есть ин-ты географии при ун-тах в Грейфсвальде, Лейпциге, Галле, Ростоке и Иене. Кроме того, при Берлинском ун-те им. А. Гумбольдта создан Ин-т политич. и экономич. географии, в Лейпциге находится Нем. ин-т страноведения. Ин-т географии Польской АН находится в Варшаве, Болгарской АН - в Софии. В Чехословакии есть Ин-т географии Словацкой АН в Братиславе и отдел экон. географии в Ин-те экономики в Праге. В Китае Ин-т географии в Пекине входит в АН КНР. В ряде социалистич. стран имеются смешанные геолого-географические или географо-экономические научные учреждения. Во Франции ин-ты географии имеются при ряде ун-тов, крупнейшие - в Париже, Страсбуре, Лилле, Бордо. Франц. Нац. ин-т географии занимается только изданием карт. В Великобритании Ин-т географии имеется в Лондонском ун-те. В ФРГ ин-ты географии также в основном находятся при ун-тах (Бонн, Рема-ген, Гёттинген, Мюнхен, Фрейбург и др.). Имеются ин-ты географии и при др. высших уч. заведениях (напр., при Мюнхенской высшей технич. школе). В Швеции ин-ты географии есть при ун-тах в Упсале, Лунде и в Стокгольме. В др. странах Зап. Европы и в США геогр. исследования обычно ведутся кафедрами географии ун-тов или геогр. об-вами. В Канаде есть геогр. отдел при правительстве, к-рый в основном ведёт картографич. работы. В Бразилии Нац. совет географии является, по существу, ин-том географии; кроме того, в Рио-де-Жанейро есть Нац. ин-т географии и статистики. В Экуадоре, Доминиканской Республике и нек-рых др. странах Лат. Америки имеются коен.-геогр. ин-ты. В Чили Ин-т географии создан при ун-те в Сантьяго. Панамериканский ин-т географии и истории находится в Мексике (г. Мехико). В Японии при Токийском университете имеется геогр. институт. Д. В. Кравченко. ГЕОГРАФИЧЕСКИЕ КАРТЫ, уменьшенные обобщённые изображения земной поверхности на плоскости, показывающие размещение, сочетания и связи природных и общественных явлений, отбираемых и характеризуемых в соответствии с назначением данной карты. Определение Г. к. только как чертежа земной поверхности недостаточно, так как Г. к. могут отображать самые разнообразные природные и социально-экономич. явления. Г. к. способны передавать пространственные изменения этих явлений во времени. Для Г. к. свойственны: особый математич. закон построения (картографические проекции), изображение явлений посредством особой знаковой системы - картографич. символов (картографич. знаков), отбор и обобщение изображаемых явлений (генерализация картографическая). Г. к. закономерно рассматривать как наглядные образно-знаковые модели. Им присущи основные черты моделей вообще: отвлечение от целого для исследования части - конкретной территории, конкретных явлений и процессов; упрощение, состоящее в отказе от учёта множества характеристик и связей и в сохранении нек-рых, наиболее существенных; обобщение, имеющее в виду выделение общих признаков и свойств, и др. Эти абстракции способствуют более глубокому познанию явлений, изображаемых на Г. к. Первая особенность Г. к.- построение при помощи картографич. проекций - позволяет получать по картам правильные данные о положении, плановых размерах и форме изображаемых земных объектов. Вторая особенность Г. к.- использование картографич. знаков как особого языка карты - даёт возможность: а) изображать земную поверхность с желательным уменьшением (т. е. в желательном масштабе), чтобы охватить единым взглядом необходимую часть или даже всю земную поверхность, воспроизводя при этом на карте те объекты, к-рые вследствие уменьшения не выражаются в масштабе карты, но по своему значению должны быть показаны; б) показывать на карте рельеф земной поверхности (напр., при помощи горизонталей), т. е. передавать неровности местности в плоском изображении; в) не ограничиваться отображением на Г. к. внешности (поверхности) предметов, а указывать их внутренние свойства (напр., на карте моря можно показать физико-химич. свойства воды, течения, рельеф и грунты морского дна и мн. др.); г) показывать распространение явлений, не воспринимаемых непосредственно нашими органами чувств (напр., магнитное склонение, аномалии силы тяжести и т. п.), и делать наглядными недоступные непосредственному восприятию связи и отношения (напр., между источниками сырья и предприятиями по его переработке); д) исключать менее значимые стороны, частности и детали, свойственные единичным объектам, и выделять их общие и существенные признаки (напр., характеризовать населённые пункты по численности населения и адм. значению, отказываясь от передачи их планировки), т. е. прибегать к абстракции. Особенно важна третья особенность Г. к.- отбор и обобщение изображаемых явлений, т. е. картографическая генерализация. Г. к. в той или иной мере используются во всех сферах человеч. деятельности. Общеизвестно их значение как путеводителей по местности. В промышленном, энергетич. и транспортном строительстве они являются основой для изысканий, проектирования и переноса в натуру инженерного проекта. В сел. хоз-ве Г. к. необходимы для землеустройства, мелиорации и вообще для учёта и наиболее рационального использования всех земельных фондов. Карты служат важным пособием для школьного и внешкольного обучения, для распространения знаний о мире и для подъёма общей культуры. Картографическая изученность территории имеет важное значение в военном деле. В условиях социалистич. строительства многие задачи нар. хоз-ва - правильная оценка географич. условий, разумное использование и восстановление ресурсов, разработка планов преобразования природы, рациональное размещение производит, сил, комплексное развитие эко-номич. районов и др.- требуют для своего решения высококачественных карт. Г. к. как средство научного исследования не только дают наглядную картину размещения явлений, но также позволяют находить закономерности этого размещения. Напр., геологич. карты, показывая геологич. строение местности, служат для выяснения закономерностей распространения месторождений полезных ископаемых. Наконец, Г. к. незаменимы для изучения пространственных взаимосвязей и развития явлений и, следовательно, могут быть средством прогноза. Картографич. изображение складывается из ряда географич. элементов, обусловливаемых темой и назначением карты. Напр., элементами содержания подробных карт местности (топографических карт) являются: воды и рельеф земной поверхности, растительный покров и грунты, населённые пункты, пути сообщения и средства связи, гос. и адм. границы и центры, а также нек-рые объекты пром-сти, сел. хоз-ва и культуры. На полях Г. к. и на свободных от картографич. изображения местах помещают вспомогательные графики и тексты, облегчающие пользование картой: легенду карты (свод картографич. знаков, применённых на карте, с необходимыми пояснениями); графики для измерения по карте расстояний, углов, площадей, координат отдельных точек, крутизны скатов и т. д.; справочные сведения о времени составления карты, об использованных источниках и т. д. Иногда на полях карты располагаются также профили, диаграммы, таблицы и текстовые данные, поясняющие и дополняющие собственно картографич. изображение. Весьма распространены общегеографич. карты, на которых гл. предметом изображения служит сама земная поверхность с объектами, на ней расположенными. Прочие карты называют тематическими. Они передают с большей полнотой и обстоятельностью какой-либо элемент (или элементы), входящий в содержание общегеографич. карты (напр., рельеф земной поверхности), или показывают явления, отсутствующие на общегеографич. картах, напр, геологич. строение местности, климатич. условия и т. п., в связи с чем различают виды тематических карт - геологич., климатич. и т. д. Тематические карты образуют два основных класса: а) карты природных явлений, или физико-географич.; б) карты общественных явлений, или социально-экономич. (населения, экономики, культуры, политико-адм., историч.). От тематич. классификации карт следует отличать их подразделение по назначению, когда из многообразия Г. к. выделяются группы специальных карт, предназначенных для определённого круга потребителей и для решения определённых задач, в частности карты учебные, туристские, навигационные, проектные и т. д. Специальными могут быть как общегеографич. (напр., туристские), так и тематич. карты (напр., учебные карты - климатич., почвенные, экономики и т. д.). Нек-рые группы спец. карт настолько специфичны, что их иногда рассматривают в качестве особого класса тематич. карт, а именно технич. карт, к к-рым относят морские навигационные карты, полётные, проектные и др. На практике широко используется классификация Г. к. по территориальному признаку (пространственному охвату), различающая карты мира в целом, карты океанов и морей, материков, их крупных частей, государств, областей, районов. Рассмотренные классификации, взятые порознь, недостаточно дифференцируют всё многообразие карт. Поэтому их часто используют совместно. Классификация по территориальному признаку обычно выбирается в качестве основной, а внутри её рубрик карты распределяются по тематике и дополнительно по назначению. Г. к. могут различаться также по широте темы. Напр., одни климатич. карты ограничивают своё содержание одним из метеорологич. элементов (температура, осадки и т. п.), другие включают несколько элементов (напр., давление воздуха и ветер), нек-рые характеризуют климат в целом. Карты узкой темы принято называть частными или отраслевыми, в данном примере частными климатич. картами, а карты, дающие полную характеристику явления, - общими, в этом примере общей климатич. картой. Многие карты показывают одновременно (совмещают) несколько явлений, каждое в своих показателях, с учётом взаимных связей явлений. Это - многоотраслевые карты; их называют комплексными. К ним принадлежат, напр., синоптические карты, показывающие совместно все основные метеорологические элементы. Г. к. неодинаковы по степени обобщения содержания. Есть карты, для к-рых используют необобщённые или мало обобщённые показатели (напр., значения метеорологич. элементов для конкретного момента времени); для других используют сильно обобщённые показатели, напр, средние месячные или даже средние годовые температуры, вычисленные по многолетним данным. Карты, выделяющие и показывающие отдельные элементы природы, населения, экономики и культуры, их свойства или особенности, являются аналитическими. Наибольшая степень обобщения наблюдается на синтетических картах, характеризующих явления как единое целое на основе соединения и совместного использования (слияния) ряда показателей. Пример - общие климатич. карты, на к-рых выделяются климатич. области по совокупности нескольких показателей (температура, осадки и др.), но без изображения этих конкретных показателей. Синтетич. карта как бы обобщает ряд частных карг. В практике встречаются карты с самыми разнообразными сочетаниями конкретных и обобщённых показателей, аналитич. и синтетич. характеристик. Напр., на многих общеэкономич. картах для пром-сти используется аналитический, а для сел. х-ва синтетич. способ картографирования. Карты, построенные по недостаточным данным, особенно когда они ставят целью истолкование наблюдённых фактов или явлений, могут иметь (в целом или в нек-рых своих частях и элементах) характер и значение гипотезы. Таковы, напр., карты различного климатич. районирования мира. Накопление новых данных позволяет производить проверку, сравнение и уточнение ранее составленных гипотетич. карт. Ценность Г. к. зависит не только от полноты, точности и современности использованных данных, но также от положенных в основу составления Г. к. научных принципов и идей, к-рые могут быть прогрессивными или устарелыми, правильными или ошибочными. Напр., В. В. Докучаев разработал для почвенных карт классификацию почв, основанную на учёте природных факторов почвообразования, и противопоставил этот взгляд неправильному представлению о почвах как о землистых горных породах, относящихся к поверхностным геологич. образованиям. Историч. справка. Простейшие картографии, рисунки, по-видимому, были известны уже в условиях первобытного общества. Древнейшие картографич. изображения, уцелевшие до наст, времени, принадлежат народам Древнего Востока (Вавилония, Египет) и Китая. При рабовладельческом строе картография достигла наивысших успехов в античное время. Греческие учёные создали первые Г. к., построенные в картографич. проекции с учётом шарообразности Земли. В средние века расцвет мореплавания (в связи с великими географич. открытиями, колонизацией Америки, торговлей с Ост-Индией и Китаем) и вызванные этим потребности навигации привели к созданию множества морских карт. Развитие картографии в эту эпоху диктовалось и образованием крупных феодально-абсолютистских гос-в, нуждавшихся в достоверных Г. к. для управления обширными территориями. В 19 в. получили широкое распространение военно-топографич. съёмки для создания подробных карт местности: топография, карты облегчали управление войсками и позволяли при боевых действиях лучше учитывать неудобства и использовать выгоды местности; позднее эти карты оказались незаменимыми при инженерных изысканиях и проектировании - дорожном, гидротехнич. и др. Дифференциация наук явилась ещё одним важным стимулом для развития картографии. Тематич. карты стали широко привлекаться для изучения размещения различных природных и общественных явлений, для исследования их пространственных закономерностей, связей и обусловленности. Потребность в тематич. картах быстро росла, когда соответствующие отрасли (напр., геология) обращались на службу практики. Значение тематич. карт ещё более увеличилось в условиях планового социалистического общества. Возможности изучения и исследования по Г. к. явлений возрастают при совместном использовании карт разной тематики. Это определяет значение и развитие картографирования комплексного, заключающегося в создании серий сопоставимых, взаимно дополняющих Г. к. и комплексных атласов. Г. к.- незаменимое по своей наглядности и лаконичности средство для хранения, передачи и получения новой информации о нашей планете и её отдельных частях - суше и океанах, об их географич. условиях и естественных богатствах, о населении, экономике, культуре и даже историч. развитии - непрерывно расширяют сферу своего действия, что влечёт за собой разработку новых видов и типов карт, а также более совершенных (в том числе автоматических) методов их создания и использования. Лит.: Салищев К. А., Картография, М., 1971; 50 лет советской геодезии и картографии, М., 1967; Тематическое картографирование в СССР, Л., 1967. К. А, Салищев. ГЕОГРАФИЧЕСКИЕ КОНГРЕССЫ международные, основная форма мировых профессиональных съездов географов. До 1-й мировой войны Г. к. созывались геогр. обществами отдельных стран, а с 20-х гг. Междунар. географич. союзом (см. Географический союз международный) и Национальными комитетами географов. Г. к. проходили: 1-й - в 1871 в Антверпене, 2-й - в 1875 в Париже, 3-й - в 1881 в Венеции, 4-й - в 1889 в Париже, 5-й - в 1891 в Берне, 6-й - в 1895 в Лондоне, 7-й - в 1899 в Берлине, 8-й - в 1904 в США (в разных городах), 9-й - в 1908 в Женеве, 10-й - в 1913 в Риме, 11-й - в 1925 в Каире, 12-й - в 1928 в Кембридже (Великобритания), 13-й - в 1931 в Париже, 14-й - в 1934 в Варшаве, 15-й - в 1938 в Амстердаме, 16-й - в 1949 в Лисабоне, 17-й - в 1952 в Вашингтоне, 18-й - в 1956 в Рио-де-Жанейро, 19-й- в 1960 в Стокгольме, 20-й - в 1964 в Лондоне, 21-й - в 1968 в Дели. Гл. задача Г. к. - обмен информацией о результатах проведённых науч. исследований и совместное обсуждение крупных геогр. проблем. Благодаря дифференциации науки основное место в программе Г. к. занимают заседания секций, где дискутируются вопросы различных отраслей географии. Тенденции к интеграции геогр. исследований выражаются в устройстве пленарных заседаний, на к-рых заслушиваются доклады по актуальным проблемам, представляющим общий интерес. В программу Г. к. включаются также собрания комиссий Междунар. географического союза и научно-популярные лекции виднейших географов мира. Во время Г. к. проводятся Генеральные ассамблеи Междунар. географического союза для решения организационных вопросов, устраиваются выставки географич. литературы и карт. Особое место в программе Г. к. принадлежит экскурсиям и симпозиумам в различных районах страны-хозяина, что позволяет участникам познакомиться со своеобразными чертами её природы, особенностями населения и хозяйства. Материалы Г. к. включают обычно тезисы сообщений, представленных на конгресс; фундаментальные монографии и сборники, отражающие уровень географии в принимающей стране; календарь конгресса, путеводители экскурсий, список членов конгресса с указанием их адресов. После конгресса публикуются труды, содержащие полные тексты всех научных докладов и обзоры дискуссий. Накопившиеся за 100 лет материалы Г. к. отражают, хотя и не в полной мере, ряд общих тенденций в эволюции геогр. исследований. На первых десяти Г. к. св. ⅓ всех выступлений посвящалось описанию путешествий в мало известные науке области земного шара, вопросам геодезии и картографии, содержанию и методам геогр. образования. Позднее секция путешествий исчезает из программы Г. к., основное внимание участников обращается на результаты углублённых исследований природной среды, естественных ресурсов, хозяйства, населения и населённых пунктов по их компонентам и региональным сочетаниям. На Г. к. возрастает количество сообщений по прикладным аспектам географии, имеющим важное экономич. значение. Географы России участвовали в работе первых Г. к. В 20-40-х гг. в связи со сложной для СССР междунар. обстановкой сов. географы были только на двух Г. к. (1931 в Париже, 1934 в Варшаве). Начиная с 1956 делегации географов СССР принимают активное участие на сессиях Г. к., где представляют сов. геогр. науку, основанную на диалектико-материалистич. методологии. В Советском Союзе подготовку к Г. к. осуществляет Национальный комитет сов. географов при Академии наук СССР. В. В. Анненков. ГЕОГРАФИЧЕСКИЕ КООРДИНАТЫ, величины, определяющие положение точки на земной поверхности: широта Ф, измеряемая углом между отвесной линией в данной точке и плоскостью земного экватора, и долгота X, измеряемая двугранным углом между плоскостью меридиана данной точки и плоскостью начального меридиана (см. рис.). Широта и долгота определяются из наблюдений небесных светил с помощью угломерных инструментов (напр., универсального инструмента, секстанта и др.), установленных с помощью уровня, и из сравнения местного времени, полученного из астрономич. наблюдений, с всемирным временем (см. Время). Г. к., так определённые, наз. астрономическими координатами точки земной поверхности. Широты отсчитываются от 0° до 90° по обе стороны от экватора, причём в Сев. полушарии Земли они считаются положительными, в Южном - отрицательными. Долготы отсчитываются от начального меридиана от 0° до 360° либо к востоку (восточная долгота), либо к западу (западная долгота; по международному счёту - положительная). Применяется также система отсчёта от 0° до 180° к востоку к западу от начала меридиана. Географические координаты точки М: широта ф (угол МСN), долгота X (угол OCN).
По международному соглашению за начальный ("первый", "нулевой") меридиан принят меридиан, проходящий через меридианный круг старой Гринвичской обсерватории в Гринвиче (Лондон) до её перевода в замок Хёрстмонсо. Прежде для этой цели служили в разное время меридианы островов Иерро (Канарские острова), Парижской и Берлинской обсерваторий и др. В России в 19 в. счёт долгот вёлся от меридиана Пулковской обсерватории. |
|
© (составление) libelli.ru 2003-2020 |
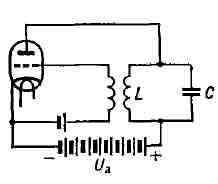
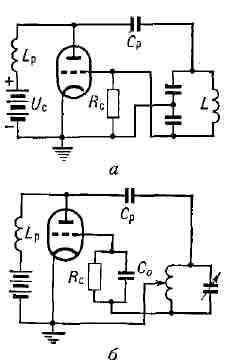
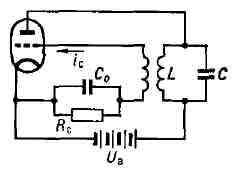
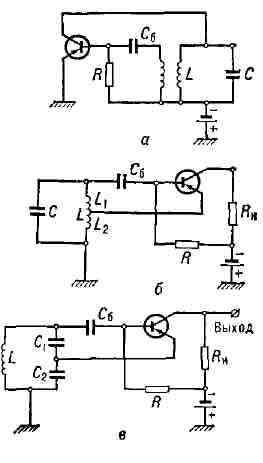
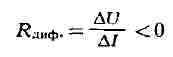 (в обычных сопротивлениях
(в обычных сопротивлениях